


网衣污损生物(亦称附着生物)是指固着或者栖息在养殖网衣部位、对人类经济活动产生不利影响的所有海洋生物的总称[1]。中国水产养殖产量多年居世界首位[2],这为民众提供了优质动物蛋白,但网衣因多缝隙、表面积大而成为污损生物的理想附着基,受其影响产生的养殖环境污染与动物病害等问题应引起广泛关注[3]。同时,随着渔业资源衰退,海洋渔业发生了由捕捞业向养殖业为主的巨大转变,1998年我国开始引进国外深水网箱技术,随后在全国沿海地区迅速发展,使深蓝渔业和渔业高质量发展也成为水产养殖可持续发展亟待解决的问题[4]。此外,养殖过程中投饵损失的饲料以及生物排泄所产生的丰富营养物质,进一步促进了养殖网衣附近水域的污损生物的生长[5]。污损生物随时间推移大量附着在网衣上而堵塞网孔,不仅影响养殖水体的交换和养殖对象的健康生长,也降低了网衣的使用年限[6]。污损生物种类组成复杂、防治困难,是生态学研究和健康养殖中面临的巨大挑战之一[7]。已有研究表明,污损生物具有种类多样、繁殖能力强、生活史短等特点,而且其生长具有规律性和阶段性[8],因此摸清污损生物的这些变化规律对网箱防污具有重要意义。
1 材料与方法
1.1 实验时间、地点
图1
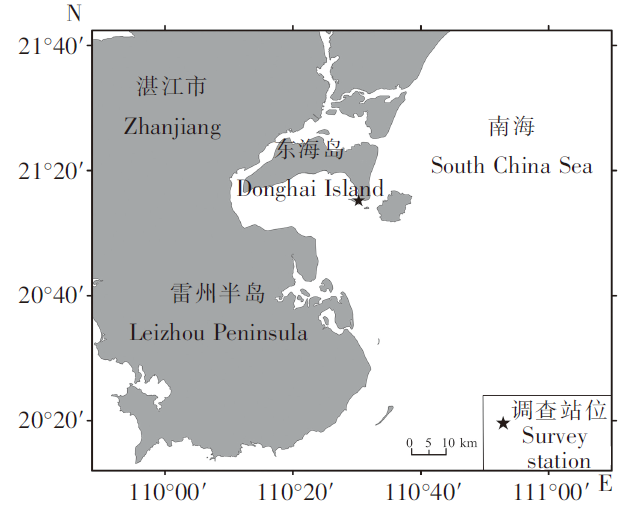
图1
雷州湾污损生物挂网实验位置
Fig.1
Location for hanging net experiment of fouling organism in Leizhou Bay
1.2 实验设计
实验网衣由网目30 mm的25 cm×25 cm聚乙烯(Polyethylene,PE)编制而成。实验共设2组,按不同的观测时间周期,每组均设季节周期网、半年周期网(上半年为12月至次年5月,下半年为6至11月)、全年周期网,分上、下两层悬挂,每个样本设4个平行样,上层距水面0.5 m,下层距水面4.5 m(图2)。网衣顶部固定于养殖平台上,底部用重锤绑缚,使网身与海底垂直。
图2
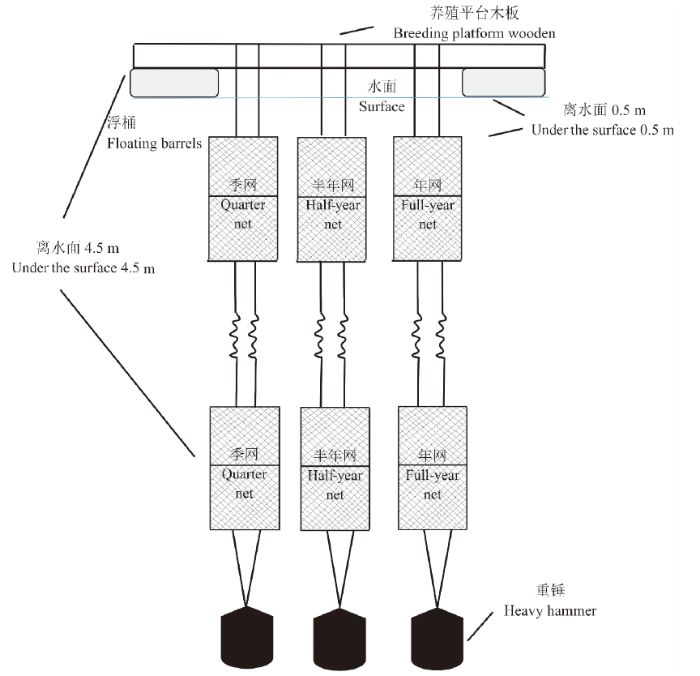
1.3 取样与鉴定
1.4 环境因子测定
每月初在挂网站点附近海域选取3个点进行环境因子检测,每次调查时间为13∶00~17∶00,由调查人员现场采用 YSI 水质检测仪测定溶解氧、pH、盐度和温度。依据GB/T 12763.6—2007《海洋调查规范 第6部分:海洋生物调查》[20]进行污损生物调查。
1.5 数据处理与分析
1.5.1 优势种
利用相对重要性指数(Index of relative importance,IRI)分析海洋污损生物群落的优势种。
式(1)中:W为某一特定物种的生物量占总生物量的百分比;N为某一特定物种的丰度占总丰度的百分比;F为某一物种的频率。IRI>25设为优势种。
海洋污损生物分为3种生活方式:运动(Motile,M)、固着(Sessiler,SE)、附着(Attached,A);按功能区分为6个功能性群组:初级生产者(Primary producers,PP)、悬浮饲养者(Suspension feeders,S)、草食动物(Herbivores,H)、食肉动物(Carnivorous,C)、杂食动物(Omnivores,O)和沉积物饲养者(Deposit feeders,D)[21]。
1.5.2 物种多样性
利用Excel对上、下水层不同时段观测周期网衣的污损生物丰度和生物量进行统计分析,群落多样性采用Shannon-Wiener 多样性指数(H')、Pielou均匀度指数(J)和Margalef丰富度指数(d)进行分析。
式(2)~(4)中:ni为第i种个体数;N为所有种个体数;S 为样品中总种类数。
1.5.3 多元统计分析
应用Excel和SPSS分别对样本进行多样性和单因素分析。应用GraphPad Prism 8.0软件对网衣污损生物群落多样性、丰富度、均匀度等指数作箱图对比。应用CANOCO 4.5软件对污损生物和水环境因子进行去趋势对应分析(Decentred correspondence analysis,DCA),当排序梯度最大值大于4.0时,选择典范对应分析(Canonical correspondence analysis,CCA)进行排序分析和作图;若在3.0~4.0之间,选择CCA或冗余分析(Redundancy analysis,RDA);若小于3.0,RDA的结果要好于CCA。
2 结果与分析
2.1 污损生物群落组成
2.1.1 种类组成
所有网衣共鉴定出污损生物35种(表1),分属11门28科35属,其中节肢动物门10种,占比28.57%;软体动物门8种,占比22.86%;扁形动物门、刺胞动物门、多孔动物门、红藻门、环节动物门、棘皮动物门、脊椎动物门、绿藻门、纽形动物门共计17种,总占比48.57%。
表1 东南码头污损生物种类组成
Tab.1
| 门 Phylum | 科 Family | 属 Genus | 种类名称 Species | 上水层 Upper water layer | 下水层 Down water layer | 春季 Spring | 夏季 Summer | 秋季 Autumn | 冬季 Winter | 上半年 First half of the year | 下半年 Second half of the year | 全年 Annual |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 节肢 动物门 Arthropoda | 地钩虾科 Podoceridae | 地钩虾属 Podocerus | 巴西地钩虾 Podocerus brasiliensis | + | + | + | + | + | + | + | + | + |
| 板钩虾科 Stenothoidae | 板钩虾属 Stenothoe | 加尔板钩虾 Stenothoe gallensis | + | + | + | + | + | + | + | + | + | |
| 蜾蠃蜚科 Corophiidae | 蜾蠃蜚属 Corophium | 莫顿蜾蠃蜚 Corophium mortonii | + | + | + | + | + | + | + | + | — | |
| 拟钩虾属 Gammaropsis | 指拟钩虾 Gammaropsis digitata | + | + | + | + | + | + | + | + | + | ||
| 藤壶科 Balanidae | 纹藤壶属 Amphibalanus | 网纹藤壶 Amphibalanus reticulatus | + | + | + | + | + | + | + | + | + | |
| 巨藤壶属 Megabalanus | 红巨藤壶 Megabalanus rosa | + | + | + | — | — | — | + | — | — | ||
| 节肢 动物门 Arthropoda | 玻璃钩虾科 Hyalidae Bulycheva | 明钩虾属 Parhyale Stebbing | 夏威夷 明钩虾 Parhyale hawaiensis | + | + | + | + | + | + | + | + | + |
| 白钩虾属 Leucothoe Leach | 翼白钩虾 Leucothoe alata | + | + | + | — | — | — | + | — | — | ||
| 梭子蟹科 Portunidae Rafinesque | 梭子蟹属 Portunus | 梭子蟹 Portunus trituberculatus | — | + | — | — | — | — | — | — | + | |
| 背尾水虱科 Anthuridae | 杯状水虱属 Cyathura | 黑斑胚筒虱 Cyathura peitrates | + | + | — | + | — | — | — | — | + | |
| 软体 动物门 Mollusca | 拟壳菜蛤科 Dreissenidae | 壳菜蛤属 Dreissena | 斑马纹贻贝 Dreissena polymorpha | + | + | + | + | + | + | + | + | + |
| 贻贝科 Mytilidae | 股贻贝属 Perna | 翡翠贻贝 Perna virids | + | + | + | + | + | — | + | + | + | |
| 贻贝属 Mytilus | 厚壳贻贝 Mytilus coruscus | + | + | — | + | + | — | + | — | + | ||
| 短齿蛤属 Brachidontes | 变化短齿蛤 Brachidontes variabilis | + | + | — | + | — | — | — | + | + | ||
| 肌蛤属 Musculus | 凸壳肌蛤 Musculus senhousei | — | + | — | — | — | — | — | — | + | ||
| 蚶科 Arcidae | 须蚶属 Barbatia | 珠肋须蚶 Barbatia yamamotoi | + | + | — | — | + | — | — | — | + | |
| 骨螺科 Muricidae | 荔枝螺属 Thais | 砺敌荔枝螺 Thais gradata | + | + | — | — | — | — | — | — | + | |
| 牡蛎科 Ostreidae | 巨牡蛎属 Crassostrea | 近江牡蛎 Crassostrea rivularis | + | + | — | — | — | — | — | — | — | |
| 刺胞 动物门 Coelenterata | 笔螅科 Pennariidae | 笔螅属 Pennaria | 笔螅 Pennaria disticha | + | + | + | + | + | — | + | + | + |
| 纵条矶 海葵科 Haliplanellidae | 纵条矶 海葵属 Haliplanella | 纵条矶 海葵 Haliplanella luciae | — | + | — | — | — | — | — | — | + | |
| 环节 动物门 Annelida | 多鳞虫科 Polynoidae | 海鳞虫属 Halosydna | 短毛海鳞虫 Halosydna nebulosa | + | + | — | + | — | — | — | + | + |
| 金扇虫科 Chrysopetalidae | 卷虫属 Bhawania | 隐头卷虫 Bhawania goodei | + | + | — | + | + | — | + | — | + | |
| 海稚虫科 Spionidae | 才女虫属 Polydora | 才女虫 Polydora sp. | + | + | — | — | — | — | + | — | + | |
| 缨鳃虫科 Sabellidae | 伪刺缨虫属 Pseudopotamilla | 伪刺缨虫 Pseudopotamilla reniformis | — | + | — | + | — | — | — | — | — | |
| 棘皮动物门 Echinodermata | 辐蛇尾科 Ophiactidae | 辐蛇尾属 Ophiactis | 近辐蛇尾 Ophiactis affinis | — | + | — | — | + | — | — | — | — |
| 多孔动物门 Porifera | 皮海绵科 Suberitidae | 荔枝海绵属 Tethya | 荔枝海绵 Tethya sp. | + | + | — | + | — | — | — | — | + |
| 纽形动物门 Nemertinea | 纵沟纽虫科 Gorgonorh ynchidae | 纵沟纽虫属 Gorgonorh ynchus | 纵沟纽虫 Gorgonorh ynchus sp. | + | + | — | — | — | — | — | + | + |
| 扁形动物门 Platyhe- lminthes | 扁虫科 Pseudoce- rotidae | 扁虫属 Pseudoceros | 外角伪涡虫 Pseudoceros exoptatus | — | + | — | — | — | — | — | — | + |
| 脊椎动物门 Verbebrata | 塘鳢科 Eleotridae | 塘鳢属 Eleotris | 塘鳢 Eleotris acanthopoma | + | — | — | + | — | — | — | — | — |
| 绿藻门 ChloropHyta | 石莼科 Ulvaceae | 浒苔属 Enteromorpha | 浒苔 Enteromorpha prolifera | + | + | + | + | + | — | + | + | + |
| 石莼属 Ulva | 石莼 Ulva lactuca | + | + | + | — | — | — | + | + | + | ||
| 礁膜科 Monos- tromataceae | 礁膜属 Monostroma | 礁膜 Monostroma | + | + | + | — | — | — | + | — | + | |
| 红藻门 RhodopHyta | 江蓠科 Ulvaceae | 江蓠属 Gracilaria | 异枝江蓠 Gracilaria bailinae | + | + | + | + | — | — | + | + | + |
| 石花菜科 Gelidiaceae | 石花菜属 Gelidium | 石花菜 Gelidium amansii | + | — | + | + | — | — | — | — | — | |
| 红翎菜科 Solieriaceae | 麒麟菜属 Eucheuma | 麒麟菜 Eucheuma muricatum | + | — | — | — | + | — | — | — | — |
注:+代表该物种于某样方出现,—则代表该物种于某样方未出现。
Notes: + meant that the species appeared in a quadrat, and — meant that the species did not appear in a quadrat.
在时间上,春季(16种)、夏季(20种)、秋季(15种)、上半年(18种)、下半年(15种)、全年(26种)均高于冬季(7种)。各个时期的网衣中,节肢动物门均有显著优势,主要包括巴西地钩虾(Podocerus brasiliensis)、加尔板钩虾(Stenothoe gallensis)、指拟钩虾(Gammaropsis digitata)等物种;其次为软体动物门,主要出现在夏、秋季,主要包括翡翠贻贝(Perna virids)、斑马纹贻贝(Dreissena polymorpha)等物种;再者为藻类,主要出现在春季,主要包括浒苔(Enteromorpha prolifera)、石莼(Ulva lactuca)等物种。
在空间上,下水层(32种)稍多于上水层(29种),污损生物种类主要包括巴西地钩虾、加尔板钩虾、指拟钩虾等物种。
2.1.2 优势种
全年网衣污损生物的优势种共有15种(表2),其中上水层12种、下水层15种。节肢动物占有绝对优势,有7种:指拟钩虾上、下水层IRI分别为4 648和3 645,网纹藤壶(Amphibalanus reticulatus)上、下水层IRI分别为3 948和2 425,加尔板钩虾上、下水层IRI分别为1 961和2 652,以及巴西地钩虾、夏威夷明钩虾、莫顿蜾蠃蜚(Corophium mortonii)、红巨藤壶(Megabalanus rosa);软体动物有4种:斑马纹贻贝、翡翠贻贝、变化短齿蛤(Brachidontes variabilis)、厚壳贻贝(Mytilus coruscus);藻类有3种:浒苔、石莼、异枝江蓠(Gracilaria bailinae);刺胞动物1种:笔螅(Pennaria disticha)。
表2 网衣污损生物优势种
Tab.2
| 水层 Water layer | 物种名称 Species | 平均密度 /ind·cm-2 Average density | 平均密度 /g·cm-2 Average density | IRI | 生活方式 Lifestyle | 功能性群体 Functional group |
|---|---|---|---|---|---|---|
| 上 Upper | 指拟钩虾 Gammaropsis digitata | 14.370 | 0.036 | 4 648 | M | D |
| 网纹藤壶 Amphibalanus reticulatus | 0.982 | 0.183 | 3 948 | SE | S | |
| 加尔板钩虾 Stenothoe gallensis | 5.470 | 0.022 | 1 961 | M | D | |
| 笔螅 Pennaria disticha | 7.700 | 0.008 | 1 845 | SE | S | |
| 巴西地钩虾 Podocerus brasiliensis | 4.404 | 0.018 | 1 576 | M | D | |
| 夏威夷明钩虾 Parhyale hawaiensis | 2.512 | 0.016 | 1 024 | M | D | |
| 斑马纹贻贝 Dreissena polymorpha | 0.346 | 0.040 | 853 | A | S | |
| 翡翠贻贝 Perna virids | 0.144 | 0.043 | 729 | A | S | |
| 莫顿蜾蠃蜚 Corophium mortonii | 1.004 | 0.005 | 323 | M | D | |
| 浒苔 Enteromorpha prolifera | 0.214 | 0.009 | 140 | A | PP | |
| 变化短齿蛤 Brachidontes variabilis | 0.176 | 0.021 | 131 | A | S | |
| 厚壳贻贝 Mytilus coruscus | 0.017 | 0.017 | 71 | A | S | |
| 下 Down | 指拟钩虾 Gammaropsis digitata | 23.120 | 0.031 6 | 3 645 | M | D |
| 加尔板钩虾 Stenothoe gallensis | 11.800 | 0.022 8 | 2 652 | M | D | |
| 网纹藤壶 Amphibalanus reticulatus | 0.740 | 0.114 0 | 2 425 | SE | S | |
| 巴西地钩虾 Podocerus brasiliensis | 4.340 | 0.021 6 | 4 473 | M | D | |
| 翡翠贻贝 Perna virids | 0.780 | 0.105 2 | 1 989 | A | S | |
| 笔螅 Pennaria disticha | 7.000 | 0.009 2 | 1 194 | SE | S | |
| 斑马纹贻贝 Dreissena polymorpha | 0.100 | 0.034 8 | 762 | A | S | |
| 莫顿蜾蠃蜚 CoropHium mortonii | 0.000 | 0.005 6 | 585 | M | D | |
| 夏威夷明钩虾 Parhyale hawaiensis | 0.260 | 0.005 6 | 273 | M | D | |
| 下 Down | 石莼 Ulva lactuca | 0.240 | 0.013 2 | 185 | A | PP |
| 红巨藤壶 Megabalanus rosa | 0.000 | 0.016 8 | 117 | SE | S | |
| 变化短齿蛤 Brachidontes variabilis | 0.740 | 0.016 0 | 103 | A | S | |
| 浒苔 EnteromorpHa prolifera | 0.340 | 0.002 8 | 40 | A | PP | |
| 厚壳贻贝 Mytilus coruscus | 0.080 | 0.018 4 | 31 | A | S | |
| 异枝江蓠 Gracilaria bailinae | 0.130 | 0.001 1 | 25 | A | PP |
注:SE.固着;A.附着;M.运动;S.悬浮喂食动物;D.沉积喂食动物;PP.初级生产者(植物)。
Notes:SE meant sessile; A meant attached;M meant motile; S meant suspension feeders; D meant deposition feeders; PP meant primary producers (plant).
本次研究优势种根据污损生物的活动和生活方式,将其分为3类:固着型、附着型和运动型,结果表明固着生物和附着生物在污损生物中占优势,再根据功能性群体分类,确定了3个功能性群体,分别为S、D、PP群。在大多数实验网衣上,S群占优势,主要代表物种为贻贝;D群是第二大优势组,主要代表物种为钩虾;而PP群主要代表物种为浒苔。根据本研究确定的优势种功能群,共有2个营养水平:PP群代表第一营养级;S和D构成第二营养级(表2)。
2.2 丰度和生物量
从空间上看,东南码头上水层的网衣污损生物丰度(37.53 ind·cm-2)显著高于下水层(29.03 ind·cm-2)(P<0.05);生物量(0.75 g·cm-2)高于下水层平均生物量(0.74 g·cm-2),但无显著性差异(P>0.05)(图3)。
图3
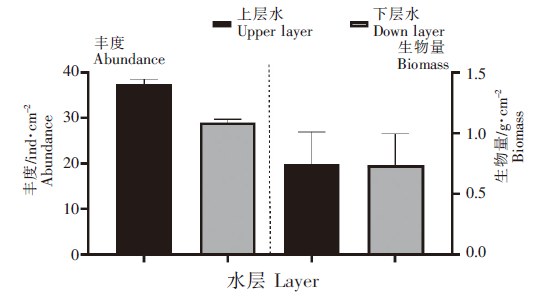
图3
不同水层网衣污损生物丰度密度和生物量密度变化
Fig.3
Changes of abundance density and biomass density of mesh fouling organisms in different layers
从时间观测周期看,网衣污损生物的丰度和生物量分布存在明显差异,上、下水层的变化趋势均表现为夏季(月份)、半年、全年高于其他时间段,冬季最低,且时间观测周期越长,丰度和生物量数值越大(图4)。
图4
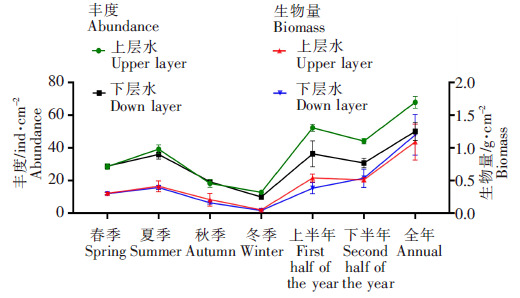
图4
不同观测周期网衣污损生物丰度密度和生物量密度变化
Fig.4
Changes of abundance density and biomass density of mesh fouling organisms in different observing periods
2.3 多样性指数
上、下水层的各个观测周期网衣多样性指数(0.985~2.010)、均匀度指数(0.539~0.852)、丰富度指数(0.539~2.856)数值相近。
网衣污损生物上、下水层7个观测周期网衣各指数的对比见图5,在空间上表现为下水层(1.149~2.010)多样性指数(H')大体上高于上水层(0.985~1.693),丰富度指数(d)(上水层:0.688~2.443;下水层:0.539~2.856)、均匀度指数(J)(上水层:0.547~0.721;下水层:0.539~0.852)无显著变化规律;在时间上表现为冬季网衣污损生物群落多样性指数(H')(春季:1.373~2.010;夏季:1.454~1.689;秋季:1.159~1.942;冬季:0.985~1.201;上半年:1.431~1.787;下半年:1.546~1.779;全年:1.423~1.751)、丰富度指数(d)(春季:1.527~1.966;夏季:1.014~1.769;秋季:1.173~1.820;冬季:0.539~1.040;上半年:1.405~1.958;下半年:1.289~1.567;全年:1.474~2.856)低于其他周期网衣,但均匀度指数
图5
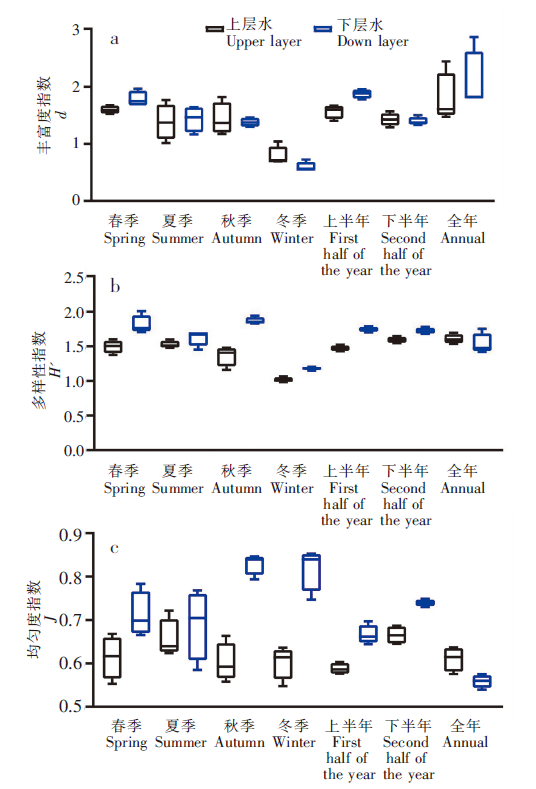
图5
不同观测周期网衣污损生物多样性变化[丰富度指数(a)、多样性指数(b)、均匀度指数(c)]
Fig.5
Variation of species diversity of net fouling organisms in different observing periods[Margalef index (a),Shannon-Wiener index(b),Pielou index(c)]
2.4 影响网衣污损生物群落分布的环境因子
对4个季节时期网衣污损生物群落丰度和生物量进行热图结合环境指标分析,结果显示春、夏季的优势种丰度和生物量较其他季节均占优势地位,其优势种丰度和生物量与温度的关系基本呈正相关。优势种节肢动物钩虾类物种的生物量和丰度显示为春、夏季占据重要优势地位,其中指拟钩虾一年四季均有分布,春季的丰度和生物量更显著;藤壶物种的丰度和生物量显示为春夏季较显著;软体动物类斑马纹贻贝和翡翠贻贝的生物量在夏、秋季较显著;藻类石莼、浒苔、石花菜(Gelidium amansii)等物种均显示为春季较显著;刺胞动物笔螅生物量和丰度显示为春夏季较显著(图6)。
图6
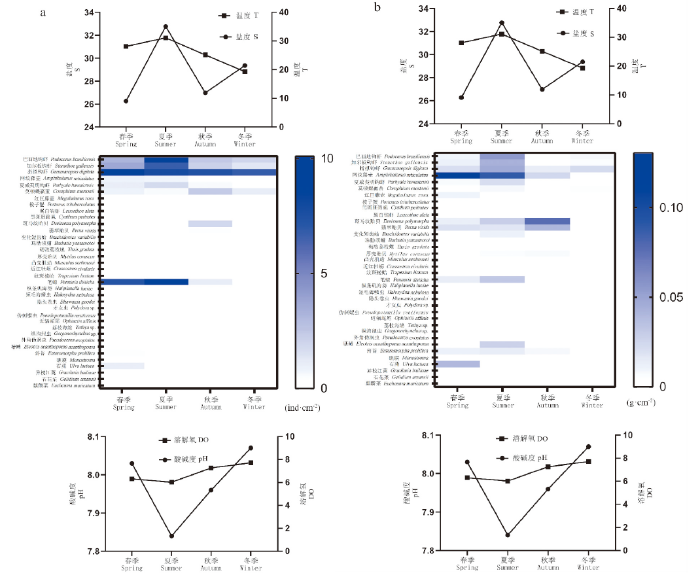
图6
不同观测季节网衣污损生物的丰度(a)和生物量(b)与环境因子的变化情况
Fig.6
Changes of the abundance (a) and biomass (b) of fouling organisms and environmental factors in different observing seasons
对4个季节观测时期网衣污损生物群落及东南码头的污损生物群落丰度和生物量进行DCA分析,排序结果显示上、下水层丰度和生物量 4 个轴最大长度分别为1.2、1.8、1.7、2.3,属于线性模型,因此再使用RDA分析环境因子对4个季节网衣污损生物群落的影响。在不同季节的上、下水层污损生物丰度、生物量-环境相关性在RDA1轴、RDA2轴相关性分别为30.89%~65.43%、11.00%~13.70%(图7)。1)上、下水层各环境因子对污损生物群落结构(种类组成丰度密度)的贡献率分别为63.6%、32.8%。分析结果均显示,盐度(上、下水层贡献率分别为40.1%、28.2%)和温度(上、下水层贡献率分别为57.4%、59.0%)为主要影响因子,盐度对夏季损生物分布影响较大,呈负相关;温度对春季、秋季、夏季的污损生物分布有极显著影响,呈正相关。2)上水层各环境因子对污损生物群落结构(种类组成生物量密度)贡献率为18.3%,分析结果均显示,盐度(贡献率为38.3%)和温度(贡献率为39.1%)为主要影响因子,温度对夏季污损生物分布有显著影响,呈正相关;盐度对夏、秋季影响较大,呈负相关。下水层各环境因子对污损生物群落结构(种类组成生物量密度)贡献率为32.7%,分析结果均显示,盐度(贡献率为21.0%)和温度(贡献率为24.3%)为主要影响因子,温度对春、夏季污损生物分布有极显著影响,呈正相关。
图7
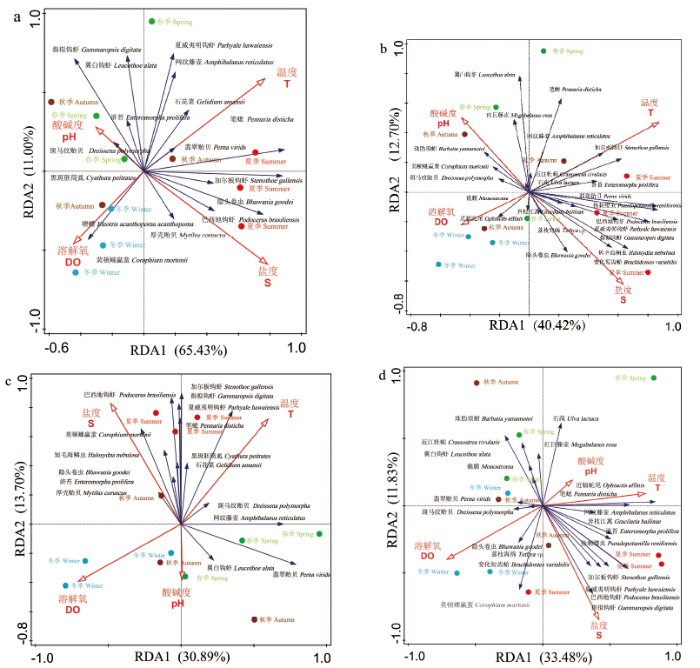
图7
不同观测时期网衣污损生物群落与环境因子RDA 分析,丰度上水层(a)、下水层(b),生物量上水层(c)、下水层(d)
Fig.7
RDA analysis of cage fouling biological community and environmental factors in different observing periods, the abundance upper water layer(a) and down water layer(b), the biomass upper water layer(c) and down water layer(d)
温度是影响污损生物群落结构的关键环境因子(平均贡献率为44.95%);盐度是影响污损生物群落结构的主要环境因子(平均贡献率为31.90%);溶解氧与网衣污损生物丰度、生物量呈负相关,但相关性较低,pH规律性不显著。
3 讨论
3.1 网衣污损生物群落组成
根据本研究结果,雷州湾网衣污损生物物种组成存在时空差异。1)从时间尺度看,节肢动物门、软体动物门及刺胞动物门在各个季节均有分布,其中节肢动物门端足目占据重要优势地位,冬季的网衣污损生物种类数较其他季节少。不同养殖海域污损生物的优势种类有很大的不同,如美国缅因州西部海洋水产养殖的软体动物贻贝为优势种[22];舟山海域水螅虫(Hydra vulgaris)、礁膜(Monostroma)、孔石莼(Ulva pertusa)、贻贝(Mytilus edulis)等为优势种[23];地中海西南部养殖场水螅和藻类为优势种[24];克罗地亚养鱼场藻类为优势种[25]。也有研究学者对海域沿岸进行污损生物现场调查,如中国东海西南沿岸海域污损生物研究调查的优势种为麦秆虫(Caprella equilibra)、中胚花筒螅(Ectopleura crocea)、海藻等[26];中国北部湾优势种为牡蛎和藤壶[27];波斯湾哈尔克岛优势种为节肢动物门和软体动物门[28]。综上结果,软体动物、节肢动物、藻类、刺胞动物出现频率高,具有显著优势,与挂网材质相关性不大,本文研究结果也符合这一生态规律。网衣污损生物附着基底为网衣,有别于船底、浮标,其中端足目因个体小在挂网早期易附着于网衣上,软体动物在幼虫阶段也易附着网衣上,并且生长迅速,在挂网后期形成体型大、生物量占优势的物种,但本研究中藻类出现频率并不为最显著,考虑其与季风气候有关,高温不利于藻类生存,但具体原因还有待进一步深入探究。2)从空间尺度看,上层29种,下层32种,表明两水层种类数相差不大,与Amini N等[28]研究指出的污损生物生物量随时间变化、不随空间变化的规律相一致。Ramadan S E等[29]对亚历山大港东部港口的4个10年(1960—1999)进行了比较研究,表明海洋污损生物多样性范围在19~35个分类单元之间,报告了El-Dekhaila港19 种海洋污染物种[30],与Ramadan S E等[29]研究结果相比,本研究的污损生物种类数接近,而与黄桂芳等[31]研究的污损生物总数154种相距甚远,但主要门类较一致,考虑本次所选取的实验材料较黄桂芳等[31]的少,具体的种类鉴定可能受表型可塑性影响,因而存在差异。综合时空生物多样性特征,本次实验表明雷州湾海域网衣污损生物生多样性高且具有一般水域的普遍特征,日后应注重对网衣的节肢动物门及软体动物门的防除探索。
3.2 网衣污损生物群落多样性
本研究还发现,从时间尺度看,春、夏、秋季的多样性指数(H')、丰富度指数(d)差异不显著,且这3个季节平均值均显著高于冬季,但均匀度指数(J)各个季节差异不显著,表明春、夏、秋季的生物多样性较冬季更为复杂。从空间尺度看,上下水层网衣污损生物群落多样性指数(H')、丰富度指数(d)、均匀度指数(J)差异不显著,表明本次取样水深0.5 m、4.5 m的污损生物生长状况差异不大。在日后深水养殖探究中,可对更深层海域做网衣污损生物多样性探究,为深水网箱养殖提供更多的参考数据。
3.3 环境因子对网衣污损生物群落分布的影响
网衣污损生物群落结构及其变化是多种环境因子在时间和空间序列上共同作用的结果,环境变化必然导致网衣污损生物群落结构和优势种的变化,而在众多环境因子中,存在起主导作用的因子[32]。本研究RDA分析结果显示,温度和盐度是影响网衣污损生物群落季节分布的主要因素,温度变化直接影响网衣污损生物的附着生长,多样性指数分析显示,春、夏、秋季较冬季更适合污损生物的生长,钩虾类和藤壶等物种生物量和丰度显示为春、夏季占据重要优势地位,斑马纹贻贝和翡翠贻贝的生物量在夏、秋季为优势种,藻类石莼、浒苔、石花菜等均显示春季较显著,这与严涛等[33]对黄渤海污损生物生态的研究结果相一致,但本文雷州湾礁膜、石莼等海藻在冬春季旺盛附着,而黄渤海礁膜、石莼等海藻一般都在温度比较高的月份旺盛附着,这可能与两地纬度差异有关。此外,Dziubińska A等[34]研究也指出污损生物群落演替受季节性影响,表明高温季节更适宜污损生物生长。RDA和丰度、生物量与环境因子的变化情况分析均显示,加尔板钩虾、笔螅、巴西地钩虾等附着生物与温度呈正相关,再进一步说明了温度对污损生物的生长响应显著。盐度是影响污损生物分布的另一关键因素,不同盐度区域污损生物的种类组成有很大的差别[35]。泥藤壶(Fistulobalanus kondakovi)适应河口低盐水域,而网纹藤壶等则在东海和南海的高盐海域出现,若水体盐度降至18甚至更低,则会妨碍网纹藤壶幼虫的发育,甚至导致其死亡[36]。在本研究中,网纹藤壶附着量较大且没有泥藤壶出现,这可能与所处海域的盐度较高密切相关;春、秋季盐度较低,受盐度和温度的复合影响,这两季的生物多样性也较为相似。
3.4 网衣污损生物管理
根据污损生物的分布特点,合理管理网衣更替,可最大限度地降低养殖成本,保证生产安全、高效。为此,建议在春、夏、秋季养殖期间提高换网频率,冬季则可适当地降低换网频率;更换的网衣应注重清理节肢动物门和软体动物门污损生物,可采用浸泡法清除节肢动物、物理冲刷法加暴晒敲打法清除软体动物,建议积极开展探索低温环保材质的网衣制作工艺,以期为绿色养殖中污损生物的防治提供参考依据。
参考文献
深水网箱清洗技术及装备研究进展
[J].
深水网箱养殖过程中,网衣会受到海洋动植物等污损生物的附着,影响网箱内的水体交换,不利于养殖鱼类的生长和存活,同时,污损生物的附着会导致网衣重量增加,容易对养殖设施造成破坏。本研究在对国内、外的网箱清洗装置及研究现状进行分析的基础上,比较了移动式水下网箱清洗装置、潮流动力型网箱清洗装置和水下清洗机器人的技术特点,并对网箱清洗技术及装备随着大数据等新兴技术发展提出了未来的发展方向。
Prevention of biofouling on aquaculture nets with ecofriendly antifouling paint formulation
[J].
南海近岸污损生物群落时空差异初步研究
[C]//
世界海洋物种登记网址
[DB/OL].(
1971.Food habits of albacore, bluefin tuna, and bonito in California waters
[J].
Successional development of fouling communities on open ocean aquaculture fish cages in the western Gulf of Maine, USA
[J].
Harmful fouling communities on fish farms in the SW Mediterranean Sea: composition, growth and reproductive periods
[J].Biological fouling organisms on fish cages represent a major issue and costly factor in marine finfish aquaculture. Cnidarians have been identified as one of the most problematical groups, contributing significantly to the occlusion and structural stress of the cage nets, but also dramatically affecting farmed species health in aquaculture facilities worldwide. Recently, significant relationships were established in different Spanish aquaculture facilities between hydrozoans and juvenile fish affected by gill injuries and mortality episodes. Community composition, growth rate and reproductive potential of biofouling were monitored on fish cages over two seasonal periods of fry cages farming, located in southern Spain (SW Alboran Sea), with a special focus on cnidarians. Biomass and community composition of biofouling changed with time and between studied periods, with a marked seasonality in colonization periods and taxonomic composition, particularly for the colonial hydrozoans. The hydroids Ectopleura larynx and Pennaria disticha were found at the highest densities. P. disticha was responsible for major biomass contribution to total hydroid biomass with the fastest growth rates. In addition, actinulae larvae of E. larynx were identified in zooplankton samples at high densities especially during periods of fry introduction in sea cages (when fish are highly vulnerable). These results corroborate evidence of the detrimental influence of fouling cnidarians in Mediterranean finfish aquaculture due to a direct harmful impact on fish health. Investigations on population dynamics, reproductive biology and envenomation potential of fouling hydrozoans should be regarded as key component of best monitoring practices to ensure good farmed fish welfare, maximization of aquaculture production and overall marine spatial planning.
The fouling of fish farm cage nets as bioindicator of aquaculture pollution in the Adriatic Sea (Croatia)
[J].
Fouling community characteristics in subtropical coastal waters of the southwestern East China Sea
[J].
Marine fouling of offshore installations in the northern Beibu Gulf of China
[J].
Spatial and temporal variations of biofouling on the oil platforms around Khark Island, Persian Gulf
[J].
Marine fouling community in the Eastern harbour of Alexandria, Egypt compared with four decades of previous studies
[J].
Factors controlling marine fouling in some Alexandria Harbours, Egypt
[J].The present work aims to relate the settlement of marine fouling organisms in three different harbours in Alexandria city which present a wide range of ecological/environmental variations. Theses are the Abu Qir, Eastern and El-Dekheila harbours. Monthly samples of marine fouling were collected from the three harbours by using white roughened polystyrene test panels (12.5x12.5 cm), as well as physicochemical, chemical and other biological data. Results were treated with multivariate statistical analysis (PCA). At the Abu Qir harbour, it was found that water temperature and salinity are the most important environmental factors controlling the temporal distribution of total fouling density, whereas water temperature and chlorophyll a are the most significant environmental factors controlling the total biomass. At the Eastern harbour it was demonstrated that ammonia and phosphate are the most significant environmental factors controlling the temporal distribution and total biomass of fouling organisms. Concerning El-Dekheila harbour, the total density and total biomass of fouling organisms were inversely correlated with the total zooplankton.
Submerged objects - a nice place to live and develop.Succession of fouling communities in the Gulf of Gdańsk, Southern Baltic
[J].
Salinity limitations on larval settlement of four barnacle species
[J].
盐度对中国东南沿海两种常见藤壶幼虫发育的影响
[J].网纹藤壶Amphibalanus reticulatus和鳞笠藤壶Tetraclita squamosa squamosa是我国东南沿海两种常见的海洋污损生物种类, 在海洋生态系统中占有重要地位。探讨这两种藤壶的幼虫发育与盐度的关系有助于丰富和发展海洋生物学知识, 并为海洋污损生物防除相关工作的开展提供数据资料。研究探讨了网纹藤壶和鳞笠藤壶无节幼虫在盐度为6‰、12‰、18‰、24‰、30‰(对照组)和36‰的培养条件下的发育状况, 观察记录5天后幼虫的存活率和各期幼虫所占百分比, 采用最小显著差数法进行差异显著性分析。结果表明, 当水体盐度≤18‰时, 会严重阻滞网纹藤壶和鳞笠藤壶幼虫的发育, 甚至导致死亡; 而当盐度≥24‰时, 这2种藤壶幼虫的成活率虽不会随盐度改变发生显著变化, 但盐度为30‰的水体更有利于网纹藤壶幼虫的发育, 而鳞笠藤壶幼虫发育状况受盐度变化的影响不大, 具备更强的耐受能力。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}