


红鳍东方鲀(Takifugu rubripes),属鲀形目(Tetraodontiformers)、鲀亚目(Tetraodontoidei)、鲀科(Tetraodontidae)、东方鲀属(Takifugu),分布于黄海、渤海、东海及北太平洋西部,作为一种有着悠久食用历史的经济鱼类[1],凭借鲜美的味道、丰富的营养及高昂的经济价值,成为中国出口创汇重要的海珍品之一[2]。目前,红鳍东方鲀的品种选育方法较为原始[3],育种进程相对鲤鱼(Cyprinus carpio)与草鱼(Ctenopharyngodon idellus)等淡水鱼较为滞后[4],且亲本群体遗传多样性有限和近交衰退,导致其养殖经济效益下降[5],因此有必要进行新品种的培育。
家系建立是水产动物种质改良的常用手段,对水产动物的遗传性状分析和育种研究具有重要作用。其中,家系选择是以在培育过程中所创建的家系作为一个选择单位,以目标筛选性状的平均值作为指标,进行选育改良。家系选育对于遗传结构改进的作用较为持续,在维持并改良水产动物经济性状等方面也可以发挥较为积极的促进作用,能够做到缓解甚至避免养殖过程中产生的经济性状衰退[6]。对鱼类的生长性能进行科学的测试在开展选育工作上有着重要的作用[7],在培育家系过程中,与生长性能相关的表型性状有生长速度、抗病力、体长、全长和体质量等,其中体质量、体长和全长是经济性性状的主要表现形式,因此可将其作为衡量亲鱼质量的重要指标。这三种生长性状也是最容易直接观测到的数量性状,它们之间的相互关系也有助于了解鱼类生长的情况[8]。在科研中,通常会对数量性状的表型值进行采集修订,然后计算各项数据的方差组分,进而对遗传参数进行估计测算[9]。
遗传力可反映被选择性状遗传能力的大小,而遗传相关则可反映出不同目标性状之间的相关程度。对这两者进行评估,有利于制定合理的选育培养,将子代的遗传增益通过选择育种达到最大化[10],如在水产养殖动物育种中,选择育种的遗传增益在生长速度上的作用超过12%[11]。遗传力在实际养殖过程中常作为重要的参考指标[12],但遗传力评估受多种因素的影响[13],因此在实际操作过程中需严格控制各项变量。于海龙等[14]测定了210、360日龄红鳍东方鲀体质量、体长和全长的遗传力;马文超等[15]则对1、2、6和18月龄的红鳍东方鲀体质量、体长和全长的生长性状进行了遗传力估计。本研究在其基础上,将测量对象限定为生长发育旺盛的红鳍东方鲀幼鱼阶段,并利用本课题组构建的红鳍东方鲀全同胞家系的生长性状测定数据,对1、2、4月龄红鳍东方鲀幼鱼进行了体质量、体长和全长数量性状的遗传力估计,旨在为红鳍东方鲀后续的育种计划提供新的数据参考。
1 材料与方法
1.1 实验材料
2022年3月,在河北省唐山牧海水产养殖有限公司挑选达到性成熟且性腺发育良好的3年龄亲本进行配对。受精的鱼卵经多次洗卵后,置于半径25 cm、深120 cm、容积约为80 L的圆锥形网箱中,并在18~20 ℃循环水中进行6~8 d的孵化。在此过程中,共构建了全同胞家系32个,选留了其中孵化成功的29个家系。家系的管理按该公司的通常方法进行:各家系分开在水泥池中喂养,水泥池规格为3 m×3 m,水深1.5 m,每池分别编号,每日进行两次吸底,循环水以30 L/h的速度进行循环,每日投喂6~7次(每隔1.5~2.0 h)。以轮虫为开口饵料;在1~2周龄后,饵料逐渐从轮虫替换为卤虫无节幼体和成虫;在1月龄以后,开始在饵料中适量添加鱼糜;在2月龄时进行分苗,同时投喂粉碎的冰鲜玉筋鱼;在4月龄时投喂6号“海童”配合饲料与鱼糜;养殖过程中尽量保证家系间各项养殖条件一致。在1、2、4月龄时,分别对体质量、体长和全长进行生长性状数据记录。
1.2 实验方法
1.2.1 生长性状的测量
分别测定29个全同胞家系1、2、4月龄幼鱼的体质量、体长和全长。每个月龄的每个家系均随机选取100尾幼鱼。
1.2.2 数据分析
1)生长性状表型值方差分析与均方构成
表1 表型值方差分析和均方构成
Tab.1
| 变异来源 Source of variation | 自由度(df) Degree of freedom | 平方和(SS) Sum of square | 均方(MS) Mean square | 期望均方[E(MS)] Expected mean square |
|---|---|---|---|---|
| 家系间 Inter the family | F-1 | SSF | MSF | n0 |
| 家系内 Within the family | N-F | SSE | MSE | |
| 总和 Summation | N-1 | SST |
本研究所构建的家系为全同胞家系,因此使用动物遗传学中全同胞相关法估计遗传力公式计算遗传力[17]。家系的加性效应值即育种值(育种值方差为
式(1)~(3)中:
2) 表型相关和遗传相关分析
本研究表型相关通过SPSS 25的Pearson系数进行计算,而遗传相关计算使用了动物遗传学中同胞协方差法:
式(4)中:X与Y分别代表两种不同的生长性状;cov为协方差;covB为组间协方差;covF为是家系间协方差;
3) 数据处理
运用Excel和SPSS 25软件对数据进行比较计算和方差分析,所得数据用平均值±标准差(
2 结果与分析
2.1 红鳍东方鲀幼鱼各家系生长性状数据分析
1月龄红鳍东方鲀幼鱼各项生长性状数据分析结果如表2所示,家系5、12、17、18在体质量、体长及全长三个指标中均有着较高的平均值以及较低的变异系数,说明这些家系在该养殖场条件下的生长表现相对稳定,对于外部环境的变化较为适应,具有较强的稳定性和预见性,因此可以将其家系亲本组合进行记录保留,作为后续的育种参考,同时从其子代中挑选合适个体进行留种,用于之后的选育工作;考虑到丰富遗传多样性,变异系数大且生长状况良好的家系1与家系19可作为后续育种的备选家系;而家系3、4、7、11、30变异系数较小,个体规格差异较小,说明其在饲养过程中具有相当程度的稳定性,但这些家系的各项生长性状的平均值相对家系5、12、17、18较低,因而适宜进行规模饲养,留待后续上市。由2月龄红鳍东方鲀幼鱼的统计测定结果(表3)可知,家系5、10、15、16、22、27生长性状较好,因此在后续选择育种中,可主要观察其生长情况,并优先考虑为育种家系;4月龄红鳍东方鲀幼鱼的统计测定结果(表4)显示,家系5在各项生长性状中具有较高的平均值且具有较低的变异系数,因此可以将其亲本组合作为日后选育的优先组合。
表2 1月龄红鳍东方鲀幼鱼各项生长性状数据分析
Tab.2
| 家系 Family | 样本数 Number of samples | 体质量/g Body mass | 体长/mm Body length | 全长/mm Total length | |||
|---|---|---|---|---|---|---|---|
| 平均值± 标准差 | 变异系数% Coefficient of variation | 平均值± 标准差 | 变异系数% Coefficient of variation | 平均值± 标准差 | 变异系数% Coefficient of variation | ||
| 1 | 100 | 0.92±0.39 | 43.06 | 26.07±3.08 | 11.83 | 30.80±4.09 | 13.27 |
| 2 | 100 | 0.85±0.28 | 32.43 | 26.97±3.37 | 12.49 | 31.97±3.99 | 12.50 |
| 3 | 100 | 0.76±0.26 | 34.63 | 25.13±2.82 | 11.24 | 30.40±3.52 | 11.58 |
| 4 | 100 | 0.79±0.25 | 32.22 | 26.30±5.07 | 19.29 | 33.10±3.79 | 11.48 |
| 5 | 100 | 1.15±0.32 | 28.16 | 28.17±3.11 | 11.04 | 34.57±3.66 | 10.60 |
| 7 | 100 | 0.81±0.22 | 26.86 | 25.57±2.39 | 9.34 | 29.43±3.02 | 10.28 |
| 8 | 100 | 1.05±0.35 | 33.69 | 27.57±2.84 | 10.31 | 32.87±4.07 | 12.37 |
| 10 | 100 | 1.41±0.45 | 31.90 | 29.47±3.28 | 11.12 | 33.83±7.46 | 22.05 |
| 11 | 100 | 0.74±0.17 | 22.43 | 24.47±1.85 | 7.57 | 28.63±2.63 | 9.19 |
| 12 | 100 | 1.04±0.26 | 25.43 | 27.77±2.61 | 9.40 | 33.30±3.72 | 11.16 |
| 13 | 100 | 1.05±0.29 | 28.46 | 26.97±3.49 | 12.94 | 33.53±4.17 | 12.43 |
| 14 | 100 | 0.77±0.26 | 34.05 | 25.20±2.92 | 11.58 | 30.37±4.23 | 13.93 |
| 15 | 100 | 1.10±0.44 | 40.38 | 29.50±4.52 | 15.34 | 35.30±5.67 | 16.06 |
| 16 | 100 | 2.15±0.54 | 25.18 | 35.30±3.66 | 10.37 | 42.97±4.26 | 9.92 |
| 17 | 100 | 1.06±0.35 | 33.45 | 27.73±2.89 | 10.46 | 33.80±3.85 | 11.38 |
| 18 | 100 | 0.97±0.22 | 23.01 | 27.70±2.69 | 9.72 | 33.10±3.38 | 10.20 |
| 19 | 100 | 0.85±0.38 | 44.22 | 25.53±3.75 | 14.68 | 31.17±5.19 | 16.66 |
| 20 | 100 | 0.98±0.28 | 28.50 | 24.37±2.55 | 10.48 | 31.03±3.42 | 11.02 |
| 21 | 100 | 0.76±0.25 | 32.67 | 24.50±5.01 | 20.44 | 31.13±3.24 | 10.42 |
| 22 | 100 | 0.78±0.30 | 38.25 | 26.07±3.89 | 14.94 | 32.13±4.85 | 15.09 |
| 23 | 100 | 0.89±0.34 | 38.44 | 27.07±3.57 | 13.19 | 33.07±4.77 | 14.43 |
| 24 | 100 | 0.62±0.22 | 34.74 | 24.57±2.72 | 11.09 | 29.40±4.06 | 13.83 |
| 25 | 100 | 0.46±0.14 | 31.06 | 22.33±4.57 | 20.48 | 28.43±3.07 | 10.80 |
| 26 | 100 | 0.51±0.16 | 30.68 | 22.53±2.62 | 11.64 | 27.70±3.28 | 11.85 |
| 27 | 100 | 0.84±0.27 | 32.19 | 25.30±3.09 | 12.20 | 31.00±3.72 | 12.01 |
| 28 | 100 | 0.68±0.18 | 27.40 | 24.80±2.73 | 11.03 | 30.57±3.48 | 11.39 |
| 29 | 100 | 0.65±0.16 | 24.15 | 21.47±4.17 | 19.45 | 27.10±2.29 | 8.47 |
| 30 | 100 | 0.51±0.16 | 30.73 | 22.20±2.47 | 11.12 | 26.60±3.17 | 11.91 |
| 32 | 100 | 0.53±0.22 | 40.38 | 23.30±3.28 | 14.08 | 28.67±4.36 | 15.20 |
表3 2月龄红鳍东方鲀幼鱼各项生长性状数据分析
Tab.3
| 家系 Family | 样本数 Number of samples | 体质量/g Body mass | 体长/mm Body length | 全长/mm Total length | |||||
|---|---|---|---|---|---|---|---|---|---|
| 平均值± 标准差 | 变异系数% Coefficient of variation | 平均值± 标准差 | 变异系数% Coefficient of variation | 平均值± 标准差 | 变异系数% Coefficient of variation | ||||
| 1 | 100 | 4.45±2.09 | 46.88 | 48.13±8.40 | 17.46 | 57.21±10.53 | 18.40 | ||
| 2 | 100 | 4.64±1.51 | 32.52 | 48.97±7.63 | 15.58 | 59.38±7.92 | 13.34 | ||
| 3 | 100 | 3.96±1.63 | 41.39 | 45.49±7.29 | 15.91 | 54.06±10.23 | 18.92 | ||
| 4 | 100 | 3.77±1.53 | 40.43 | 45.49±8.22 | 18.06 | 56.62±8.86 | 15.64 | ||
| 5 | 100 | 5.81±2.47 | 42.58 | 50.44±8.44 | 16.73 | 61.58±10.77 | 17.49 | ||
| 7 | 100 | 6.39±2.23 | 34.97 | 54.50±7.04 | 12.91 | 65.29±8.89 | 13.62 | ||
| 8 | 100 | 6.52±2.66 | 40.88 | 52.33±8.47 | 16.18 | 63.99±11.29 | 17.65 | ||
| 10 | 100 | 7.62±2.56 | 33.58 | 56.91±8.02 | 14.09 | 68.63±9.49 | 13.82 | ||
| 11 | 100 | 5.68±1.97 | 34.75 | 51.68±6.78 | 13.12 | 63.06±9.26 | 14.68 | ||
| 12 | 100 | 6.30±2.19 | 34.74 | 54.41±7.26 | 13.34 | 66.35±9.57 | 14.42 | ||
| 13 | 100 | 6.01±2.38 | 39.76 | 52.47±7.90 | 15.06 | 64.58±10.49 | 16.24 | ||
| 14 | 100 | 5.88±1.81 | 30.72 | 52.14±7.85 | 15.05 | 65.10±7.76 | 11.92 | ||
| 15 | 100 | 6.82±2.38 | 34.88 | 56.73±7.47 | 13.18 | 69.27±8.69 | 12.53 | ||
| 16 | 100 | 10.74±2.77 | 25.75 | 64.61±6.27 | 9.71 | 80.47±7.54 | 9.37 | ||
| 17 | 100 | 5.02±1.69 | 33.72 | 49.92±5.86 | 11.73 | 61.68±7.59 | 12.31 | ||
| 18 | 100 | 5.59±2.13 | 38.10 | 52.74±7.52 | 14.25 | 64.40±9.29 | 14.44 | ||
| 19 | 100 | 5.34±2.09 | 39.34 | 50.82±7.04 | 13.86 | 62.18±8.43 | 13.56 | ||
| 20 | 100 | 4.99±1.48 | 29.61 | 47.21±5.41 | 11.47 | 59.19±7.05 | 11.92 | ||
| 21 | 100 | 4.96±1.98 | 40.10 | 48.61±8.86 | 18.23 | 60.02±9.15 | 15.25 | ||
| 22 | 100 | 6.86±3.31 | 48.21 | 55.93±10.56 | 18.89 | 68.99±12.57 | 18.21 | ||
| 23 | 100 | 6.28±2.22 | 35.41 | 56.17±7.20 | 12.82 | 68.96±9.12 | 13.23 | ||
| 24 | 100 | 6.31±2.26 | 35.80 | 55.59±7.21 | 12.97 | 68.53±9.01 | 13.15 | ||
| 25 | 100 | 4.99±1.59 | 32.00 | 51.36±6.44 | 12.54 | 63.00±7.50 | 11.91 | ||
| 26 | 100 | 5.06±1.68 | 33.29 | 50.24±6.56 | 13.05 | 61.51±7.65 | 12.43 | ||
| 27 | 100 | 6.93±2.53 | 36.51 | 55.10±7.82 | 14.19 | 68.21±9.64 | 14.13 | ||
| 28 | 100 | 6.13±2.08 | 33.97 | 52.33±7.04 | 13.45 | 64.19±8.39 | 13.07 | ||
| 29 | 100 | 5.00±2.36 | 47.26 | 46.90±8.85 | 18.87 | 57.90±10.95 | 18.92 | ||
| 30 | 100 | 6.18±2.08 | 33.66 | 54.00±6.92 | 12.82 | 65.50±8.77 | 13.40 | ||
| 32 | 100 | 5.49±1.82 | 33.23 | 53.20±7.31 | 13.74 | 65.51±8.57 | 13.07 | ||
表4 4月龄红鳍东方鲀幼鱼各项生长性状数据分析
Tab.4
| 家系 Family | 样本数 Number of samples | 体质量/g Body mass | 体长/mm Body length | 全长/mm Total length | ||||
|---|---|---|---|---|---|---|---|---|
| 平均值± 标准差 | 变异系数% Coefficient of variation | 平均值± 标准差 | 变异系数% Coefficient of variation | 平均值± 标准差 | 变异系数% Coefficient of variation | |||
| 1 | 100 | 76.47±9.30 | 12.16 | 137.33±5.35 | 3.90 | 157.49±8.01 | 5.09 | |
| 2 | 100 | 67.83±8.75 | 12.90 | 132.41±5.68 | 4.29 | 147.04±6.94 | 4.72 | |
| 3 | 100 | 67.75±14.10 | 20.82 | 127.29±6.77 | 5.32 | 141.84±8.32 | 5.87 | |
| 4 | 100 | 66.14±11.11 | 16.81 | 127.88±6.42 | 5.02 | 144.46±7.52 | 5.2 | |
| 5 | 100 | 73.24±14.40 | 19.66 | 128.92±8.56 | 6.64 | 148.11±10.78 | 7.28 | |
| 7 | 100 | 61.54±8.52 | 13.85 | 125.52±5.91 | 4.71 | 140.43±6.77 | 4.82 | |
| 8 | 100 | 62.05±11.04 | 17.80 | 123.89±6.73 | 5.43 | 140.03±8.75 | 6.25 | |
| 10 | 100 | 80.82±14.30 | 17.70 | 133.74±7.42 | 5.55 | 150.06±8.98 | 5.98 | |
| 11 | 100 | 62.03±10.67 | 17.21 | 126.09±6.87 | 5.44 | 142.47±9.14 | 6.42 | |
| 12 | 100 | 70.32±11.95 | 16.99 | 129.80±7.37 | 5.68 | 144.68±8.89 | 6.15 | |
| 13 | 100 | 77.58±10.37 | 13.37 | 133.51±6.78 | 5.07 | 150.93±8.09 | 5.36 | |
| 14 | 100 | 60.36±11.47 | 19.00 | 127.05±6.81 | 5.36 | 145.52±8.56 | 5.88 | |
| 15 | 100 | 75.87±12.12 | 15.97 | 133.32±6.69 | 5.05 | 148.83±8.43 | 5.66 | |
| 16 | 100 | 71.47±9.88 | 13.82 | 134.56±5.67 | 4.22 | 153.76±7.28 | 4.73 | |
| 17 | 100 | 66.28±9.26 | 13.98 | 127.75±5.83 | 4.56 | 143.48±6.79 | 4.74 | |
| 18 | 100 | 67.23±11.26 | 16.74 | 130.88±7.03 | 5.37 | 146.67±7.87 | 5.37 | |
| 19 | 100 | 75.11±10.10 | 13.44 | 132.86±5.75 | 4.33 | 151.35±8.87 | 5.86 | |
| 20 | 100 | 62.47±10.01 | 16.04 | 122.09±6.48 | 5.31 | 139.40±7.89 | 5.66 | |
| 21 | 100 | 77.87±10.02 | 12.87 | 138.05±5.61 | 4.07 | 154.73±7.34 | 4.75 | |
| 22 | 100 | 78.69±14.28 | 18.15 | 138.16±7.49 | 5.42 | 158.39±8.32 | 5.25 | |
| 23 | 100 | 66.36±10.90 | 16.43 | 131.04±6.86 | 5.23 | 149.66±8.53 | 5.70 | |
| 24 | 100 | 67.48±10.28 | 15.23 | 130.17±6.33 | 4.86 | 147.95±8.23 | 5.57 | |
| 25 | 100 | 59.75±9.21 | 15.41 | 123.03±6.19 | 5.04 | 138.48±8.42 | 6.08 | |
| 26 | 100 | 63.24±8.41 | 13.30 | 126.30±5.74 | 4.54 | 141.74±6.76 | 4.77 | |
| 27 | 100 | 71.31±10.05 | 14.09 | 131.55±5.98 | 4.55 | 151.07±7.94 | 5.26 | |
| 28 | 100 | 73.63±10.16 | 13.80 | 131.34±6.69 | 5.10 | 153.37±8.72 | 5.68 | |
| 29 | 100 | 65.94±8.38 | 12.70 | 127.16±6.03 | 4.74 | 144.91±7.15 | 4.93 | |
| 30 | 100 | 66.53±10.71 | 16.10 | 132.08±6.47 | 4.90 | 147.66±7.57 | 5.13 | |
| 32 | 100 | 70.17±9.56 | 13.63 | 132.78±6.02 | 4.53 | 148.15±6.99 | 4.72 | |
2.2 各月龄红鳍东方鲀幼鱼群体生长性状数据的单因素方差分析
表5为各月龄红鳍东方鲀幼鱼生长性状数据单因素方差分析结果,经F检验,各个生长性状差异均表现为极显著(P<0.01),由此可认为本次计算所得出的结果可信度较高。
表5 各月龄红鳍东方鲀生长性状数据单因素方差分析
Tab.5
| 月龄 Month old | 性状 Character | 因素 Factor | 平方和(SS) Sumof square | 自由度(df) Degree of freedom | 均方(MS) Mean square | F | P值 P value | |||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 体质量 | 家系间 | 89.01 | 28 | 3.07 | 34.67 | 0 | |||
| 家系内 | 77.01 | 2 899 | 0.09 | 0 | ||||||
| 体长 | 家系间 | 6 321.63 | 28 | 217.99 | 19.21 | 0 | ||||
| 家系内 | 9 872.33 | 2 899 | 11.35 | 0 | ||||||
| 体全长 | 家系间 | 8 452.96 | 28 | 291.48 | 17.61 | 0 | ||||
| 家系内 | 14 402.60 | 2 899 | 16.56 | 0 | ||||||
| 2 | 体质量 | 家系间 | 5 234.56 | 28 | 180.50 | 39.19 | 0 | |||
| 家系内 | 13 680.31 | 2 899 | 4.61 | 0 | ||||||
| 体长 | 家系间 | 50 734.68 | 28 | 1 749.47 | 31.14 | 0 | ||||
| 家系内 | 166 858.28 | 2 899 | 56.18 | 0 | ||||||
| 体全长 | 家系间 | 81 490.79 | 28 | 2 810.03 | 33.69 | 0 | ||||
| 家系内 | 247 735.13 | 2 899 | 83.41 | 0 | ||||||
| 4 | 体质量 | 家系间 | 99 592.53 | 28 | 3 556.88 | 29.95 | 0 | |||
| 家系内 | 340 945.67 | 2 899 | 118.76 | 0 | ||||||
| 体长 | 家系间 | 50 106.91 | 28 | 1 789.53 | 42.31 | 0 | ||||
| 家系内 | 121 421.87 | 2 899 | 42.29 | 0 | ||||||
| 体全长 | 家系间 | 78 499.86 | 28 | 2 803.57 | 42.61 | 0 | ||||
| 家系内 | 188 922.36 | 2 899 | 65.80 | 0 | ||||||
2.3 各月龄红鳍东方鲀幼鱼遗传力计算
各月龄红鳍东方鲀幼鱼生长性状方差分析和遗传力估计结果见表6。由表6可知,1月龄红鳍东方鲀幼鱼体质量、体长以及全长的遗传力分别为0.54、0.39、0.36,加性效应方差分别为0.103、7.123、9.480,表型方差分别为0.192、18.471、26.035,残差分别为0.089、11.348、16.555;2月龄红鳍东方鲀幼鱼分别为0.28、0.23、0.25,加性效应方差分别为1.777、17.104、27.542,表型方差分别为6.383、73.285、110.955,残差分别为4.606、56.181、83.413;4月龄红鳍东方鲀幼鱼分别为0.23、0.29、0.29,加性效应方差分别为34.728、17.649、27.654,表型方差分别为153.483、59.942、55.485,残差分别为118.755、42.293、65.804。这三种数量性状的遗传力变化趋势如图1所示,体质量遗传力随着幼鱼月龄的增大而逐渐降低,体长及全长遗传力随着幼鱼月龄的增大而降低后又略有回升。整体上,红鳍东方鲀幼鱼的遗传力随着月龄的增加而逐渐降低。
表6 各月龄红鳍东方鲀生长性状方差分析和遗传力估计
Tab.6
| 月龄 Month old | 性状 Character | 加性效应方差 Additive effect variance | 表型方差 Phenotypic variance | 残差 Residual error | 遗传力h2 Heritability | 标准误σh Standard error |
|---|---|---|---|---|---|---|
| 体质量 | 0.103 | 0.192 | 0.089 | 0.54 | 0.14 | |
| 1 | 体长 | 7.123 | 18.471 | 11.348 | 0.39 | 0.14 |
| 全长 | 9.480 | 26.035 | 16.555 | 0.36 | 0.17 | |
| 体质量 | 1.777 | 6.383 | 4.606 | 0.28 | 0.05 | |
| 2 | 体长 | 17.104 | 73.285 | 56.181 | 0.23 | 0.16 |
| 全长 | 27.542 | 110.955 | 83.413 | 0.25 | 0.19 | |
| 体质量 | 34.728 | 153.483 | 118.755 | 0.23 | 0.23 | |
| 4 | 体长 | 17.649 | 59.942 | 42.293 | 0.29 | 0.14 |
| 全长 | 27.654 | 55.485 | 65.804 | 0.29 | 0.18 |
图1
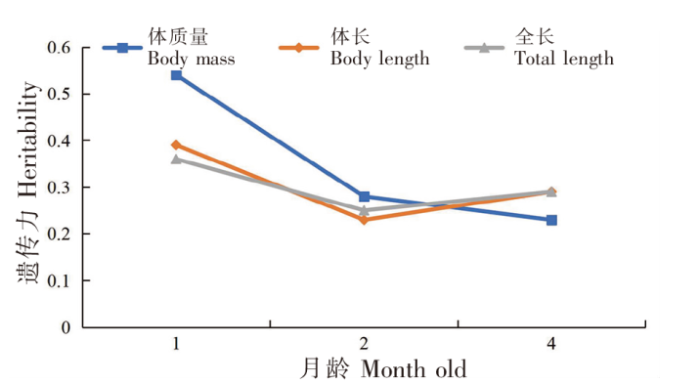
图1
红鳍东方鲀生长性状遗传力随月龄变化的情况
Fig.1
Line gram of growth traits with monthly age in T. rubripes
2.4 各月龄幼鱼性状间的遗传相关和表型相关
表7为各月龄红鳍东方鲀幼鱼体质量、体长、全长数量性状间的遗传相关和表型相关分析结果。1月龄红鳍东方鲀幼鱼体长、体质量间的遗传相关为0.889,表型相关为0.851;体质量、全长间的遗传相关为0.873,表型相关为0.864;体长、全长间的遗传相关为0.947,表型相关为0.894,遗传相关均表现为高度相关(0.8~1.0属于高度相关[15]);2月龄体长、体质量间的遗传相关为0.828,表型相关为0.917;体质量、全长间的遗传相关为0.820,表型相关为0.932;体长、全长间的遗传相关为0.897,表型相关为0.952,遗传相关均表现为高度相关;4月龄体长、体质量间的遗传相关为0.769,表型相关为0.779;体质量、全长间的遗传相关为0.726,表型相关为0.801;体长、全长间的遗传相关为0.912,表型相关为0.941,其中,体长、体质量间的遗传相关表现为显著相关(0.5~0.8属于显著相关[15]),其余遗传相关表现为高度相关,且3个月龄的红鳍东方鲀幼鱼各生长性状间的表型相关结果均达到极显著水平(P<0.01)。
表7 红鳍东方鲀各月龄各性状间遗传相关与表型相关
Tab.7
| 月龄 Month old | 性状Character | 体质量Body mass | 体长Body length | 全长 Total length |
|---|---|---|---|---|
| 1 | 体质量 | 1.000 | 0.889 | 0.873 |
| 体长 | 0.851** | 1.000 | 0.947 | |
| 全长 | 0.864** | 0.894** | 1.000 | |
| 2 | 体质量 | 1.000 | 0.828 | 0.820 |
| 体长 | 0.917** | 1.000 | 0.897 | |
| 全长 | 0.932** | 0.952** | 1.000 | |
| 4 | 体质量 | 1.000 | 0.769 | 0.726 |
| 体长 | 0.779** | 1.000 | 0.912 | |
| 全长 | 0.801** | 0.941** | 1.000 |
注:上三角为遗传相关,下三角为表型相关(**表示在0.01水平上显著,双尾)。
Notes:The upper triangle was genetically related and the lower triangle was phenotypically related (** indicated significant at the 0.01 level,two-tailed).
3 讨论
3.1 红鳍东方鲀实验月龄的选择
通过测定样本的生长数据,可以估计出它们的早期遗传力,实现养殖优化。在实际生产过程中,生长速度是被高度重视的指标[18],而红鳍东方鲀的养殖周期较长,且性成熟时期较晚,雌、雄个体均需3年龄才能达到性成熟[19],如不筛选出性状优良的个体则会造成不必要的养殖成本浪费,因此有必要在其早期阶段进行性状筛选[20]。同时,早期选择存在着相关性优势,如Gall G等[21]在对罗非鱼(Oreochromis mossambicus)改良育种的研究中,认为通过体长对体质量的间接选育应在幼鱼阶段进行。而目前关于红鳍东方鲀早期阶段的遗传力估计报道相对较少,因此本研究选取的样本均为生长早期阶段的红鳍东方鲀幼鱼。此外,本研究选取的3个月龄均是养殖过程中需要注意的重要时间节点,1、2月龄是苗种交易及进行分流的时刻,4月龄则是剪除牙齿及选取个体进行PIT标记的时刻,因此选择该时间节点进行遗传力估计,有助于进行选择育种[15]。
3.2 红鳍东方鲀遗传力的划分及其影响因素
关于鱼类生长性状遗传力的报道情况有很多,如唐瞻杨等[24]对3、4、5月龄的尼罗罗非鱼(Oreochromis niloticus)进行了体质量、全长、体长、头长、躯干长、体高、尾柄长、尾柄高及体宽9个数量性状的遗传力估计;Withler R E 等[25]估计了银鲑(Oncorhynchus kisutch)18月龄的体质量遗传力;Blanc J M等[26]估计了褐鳟(Salmo trutta)6月龄的体质量与体长遗传力;关于鳕鱼(Gadus morhua)[27]、山女鳟(Oncorhynchus masou masou)[28]、全州禾花鲤(Cyprinus carpiovar.Quanzhouensis)[29]、金鲷(Sparus aurata)[30]以及牙鲆(Paralichthys olivaceus)[31]等鱼类的各项生长性状遗传力估计也有报道。本研究中,红鳍东方鲀的生长性状遗传力在1月龄时达到峰值,之后随着鱼体月龄的增大而逐渐降低,但4月龄的体长和全长遗传力相较于2月龄有了细微程度上升,这与尼罗罗非鱼体质量遗传力变化的整体趋势[24]相同,其在3月龄为0.66,在4月龄时下降到0.17,而5月龄上升为0.25。出现这种情况最有可能的原因是父本效应[9]和母本效应[32],但本研究所使用的方法无法考虑到两种效应产生的作用。
本研究估计了红鳍东方鲀幼鱼阶段的体质量、体长以及全长的遗传力,而生物的生长性状受到基因与环境的影响较大。本研究中外部条件控制相对得当,但养殖密度仍然成为影响红鳍东方鲀遗传力的重要因素[33],这种情况在估计脊尾白虾(Exopalaemon carinicauda)[9]遗传力的过程中也有出现。红鳍东方鲀存在咬尾及互相残杀的天性,这会直接导致其随着时间的推移而养殖密度发生不同程度的改变。有研究表明,在红鳍东方鲀孵出的第6天后,便开始出现互相残杀[34]的现象,但本研究持续时间较长,导致红鳍东方鲀幼苗之间的互相残杀在一定程度上改变了养殖密度,从而造成数据偏差,如在后续跟进家系16的过程中,发现其与其他家系相比存在一定程度的密度降低,因而其体质量、体长以及全长的平均值较高。此外,红鳍东方鲀还存在钻沙潜底的天性[34],虽然在研究过程中已对其进行实时观测并稳定动态密度,但保持各个家系的密度相同仍具有一定的难度,这些问题需留待后续研究解决。
4 结论
本研究构建了29个红鳍东方鲀全同胞家系,并估计了红鳍东方鲀幼鱼阶段1、2、4月龄的体质量、体长、全长3个数量性状的遗传力,其中1月龄红鳍东方鲀幼鱼的体质量、体长以及全长遗传力分别为0.54、0.39、0.36,均表现为高遗传力;2月龄红鳍东方鲀幼鱼的体质量、体长以及全长遗传力分别为0.28、0.23、0.25,均表现为中等遗传力;4月龄红鳍东方鲀幼鱼的体质量、体长以及全长遗传力分别为0.23、0.29、0.29,均表现为中等遗传力。在选种关键阶段的1、2月龄时期,所有性状数据在表型相关与遗传相关上均达到0.8以上,表现为高度正相关。本研究获得的红鳍东方鲀在早期幼鱼阶段的体质量、体长及全长的生长性状遗传参数,可为红鳍东方鲀的继续选育工作提供参考数据。
参考文献
Experimental infection of several fish species with the causative agent of Kuchijirosho (snout ulcer disease) derived from the tiger puffer Takifugu rubripes
[J].
Kuchijirosho (snout ulcer disease) is a fatal epidemic disease which affects the tiger puffer, Takifugu rubripes, a commercial fish species in Japan and Korea. To assess the possibility that non-tiger puffer fish can serve as reservoirs of infection, 5 fish species were challenged by infection with the extracts of Kuchijirosho-affected brains from cultured tiger puffer: grass puffer T. niphobles, fine-patterned puffer T. poecilonotus, panther puffer T. pardalis, red sea bream Pagrus major, and black rockfish Sebastes schlegeli. When slightly irritated, all these species, especially the puffer fish, exhibited typical signs of Kuchijirosho, i.e., erratic swimming, biting together and bellying out (swelling of belly), as generally observed in tiger puffers affected by Kuchijirosho. Although the mortalities of the 2 non-puffer species were lower, injection of the extracts prepared from the brains of both inoculated fish into tiger puffer resulted in death, indicating that the inoculated fish used in this experiment have the potential to be infected with the Kuchijirosho agent. Condensations of nuclei or chromatin in the large nerve cells, which is a major characteristic of Kuchijirosho, were histopathologically observed to some extent in the brains of all kinds of puffer fish species infected. These findings suggest that the virus can spread horizontally among wild and cultured puffers and even among fishes belonging to different orders.
鱼类新品种(系)生长性能测试工作探讨
[J].生长性能是鱼类新品种(系)选育的重要性状,而影响鱼类生长性能的因子主要有鱼类的基因、养殖环境、养殖活动及有意或无意的人为干扰。1996—2021年我国被认定的鱼类养殖品种有120个,但由于未建立相对统一的测试技术评价标准或规范,仍有部分被认定品种未能应用于生产。为此,通过介绍鱼类品种(系)的认定和测试技术现状,重点分析生长性能测试评价中的主要影响因素,提出建立测试基地和试验平台、完善试验设计、加强养殖管理、减少人为干扰、科学采集数据、结果评价及其判定等一整套的技术建议,为我国开展鱼类新品种(系)的测试场建设,科学、规范、公正地进行鱼类新品种(系)测试和评价提供可行的技术参考,也可为其他水产养殖新品种(系)测试场的建设和测试工作提供借鉴。
黄鳍棘鲷全长遗传力估计
[J].黄鳍棘鲷是我国东南沿海重要的海水经济鱼类,是目前尚没有通过国家原良种审定委员会审定的新品种。为做好黄鳍棘鲷新品种选育工作,培育出生长速度快的新品种,需估计生长性状的遗传力。本研究以黄鳍棘鲷为研究对象,采用巢式设计和人工授精方法,构建了19个黄鳍棘鲷全同胞家系(包括7个半同胞家系),测量35日龄黄鳍棘鲷的生长数据,并使用MTDFREML软件对其全长进行遗传力估计。结果表明,35日龄黄鳍棘鲷全长的遗传力为(0.234±0.047),属于中等水平遗传力。研究结果可为黄鳍棘鲷的良种选育提供参考资料。
Climate mismatch between introduced biological control agents and their invasive host plants:improving biological control of tropical weeds in temperate regions
[J].Many weed biological control programs suffer from large-scale spatial variation in success due to restricted distributions or abundances of agents in temperate climates. For some of the world’s worst aquatic weeds, agents are established but overwintering conditions limit their survival in higher latitudes or elevations. The resulting need is for new or improved site- or region-specific biological control tools. Here, we review this challenge with a focus on low-temperature limitations of agents and propose a roadmap for improving success. Investigations across spatial scales, from global (e.g., foreign exploration), to local (selective breeding), to individual organisms (molecular modification), are discussed. A combination of traditional (foreign) and non-traditional (introduced range) exploration may lead to the discovery and development of better-adapted agent genotypes. A multivariate approach using ecologically relevant metrics to quantify and compare cold tolerance among agent populations is likely required. These data can be used to inform environmental niche modeling combined with mechanistic modeling of species’ fundamental climate niches and life histories to predict where, when, and at what abundance agents will occur. Finally, synthetic and systems biology approaches in conjunction with advanced modern genomics, gene silencing and gene editing technologies may be used to identify and alter the expression of genes enhancing cold tolerance, but this technology in the context of weed biological control has not been fully explored.
An 8-year breeding program for Asian seabass lates calcarifer:genetic evaluation,experiences,and challenges
[J].Selective breeding for marine finfish is challenging due to difficulties in reproduction, larval rearing, and on-growth in captive environments. The farming of Asian seabass (Lates calcarifer) has all these problems and our knowledge of the quantitative genetic information (heritability and correlations) of traits necessary for commercial exploitation is poor. The present study was conducted to address this knowledge gap and to provide information that can be applied to sea bass and other aquaculture species. We carried out a comprehensive genetic evaluation for three traits (body weight, total length, and survival) collected from a breeding population for Asian seabass over an eight-year period from 2010 to 2017. Statistical analysis was carried out on 4,567 adult fish at 105, 180, 270, 360, 450, and 570 days post-hatch (dph). The heritabilities (h(2)) estimated for body weight and length using linear mixed model were moderate to high (0.12 to 0.78 and 0.41 to 0.85, respectively) and they differed between the measurement periods. Survival during grow-out phase was analyzed using threshold logistic and probit models. The heritability estimates for survival rate on the underlying liability scale (h(L)(2)) varied from 0.05 to 0.21. When the observed heritability obtained from the linear mixed model was back-transformed to the liability scale, they were similar but not significant. In addition, we examined effects of genotype by environment (GxE) interaction on body traits. The genetic correlation for body weight between tank and sea cage cultures were high (0.91-0.94) in the first and second rearing periods (180 and 270 dph) but the correlation was decreased to 0.59 +/- 0.33 at 360 dph. This suggests that the genotype by environment interaction is important for body traits in this population. Furthermore, the genetic correlations of body weights between different measurement periods were moderate but different from one. This suggests that body weights measured at different time points may be different traits and selection for improved early weight may not capture all genetic expressions in subsequent rearing periods in Asian seabass. Selection of the nucleus in sea cages may produce genotypes that do not perform equally well in tanks, although this deserves further studies to determine a suitable selection environment and optimize the breeding program. This paper discusses challenges encountered during implementation of the selection program for L. calcarifer.
The endocrine regulation network of growth hormone synthesis and secretion in fish:emphasis on the signal integration in somatotropes
[J].
Application of mixed-model techniques to fish breed improvement:analysis of breeding value selection to increase 98-day body weight in tilapia
[J].
大刺鳅18月龄体质量的育种值和遗传力估计
[J].随着人工养殖大刺鳅产业的逐渐兴起,苗种种质混乱和遗传背景不清晰等问题日益突出,不利于该品种的商业化开发。为探究大刺鳅的选育潜力,本研究以野生大刺鳅为亲本,通过全同胞育种模式构建了17个全同胞家系。经过18个月龄的养殖,最终获得7个有效家系,并采用单性状动物模型,以各家系亲本的体质量为协变量,估算了18月龄体质量的遗传力和个体BLUP育种值。通过18月龄大刺鳅全同胞家系体质量的方差组分评估结果显示,遗传力达到了(0.468 1±0.173 7),t检验P < 0.01。此外,体质量与其育种值之间呈正相关线性关系,雄鱼回归方程为y= 0.393 6x - 84.193 0(R2 = 0.755 4,P < 0.01);雌鱼回归方程为y = 0.517x - 64.126 (R2 = 0.817 8,P < 0.01)。个体育种值前50的雌性个体主要集中在3个家系中(B、E和D),尤其是家系B,雌性育种值前24和雄性育种值前19均来源于该家系。其中,雄鱼最大育种值为75.09,雌鱼最大育种值为67.49,均来自于家系B。本研究显示大刺鳅体质量具有较高的遗传力,可以通过表型或BLUP育种值的方式进行选育。
Genetic variation in body weight and flesh colour of the coho salmon (Oncorhynchus kisutch) in British Columbia
[J].
Genetic variation of body size,condition and pyloric caeca number in juvenile brown trout,Salmo trutta L.
[J].
Genetic variation and genotype by location interaction in body weight,spinal deformity and sexual maturity in Atlantic cod (Gadus morhua) reared at different locations off Norway
[J].
Heritability of body weight and fork length for Oncorhynchus masou masou
[J].
Estimates of heritability and genetic correlation for body length and resistance to fish pasteurellosis in the gilthead sea bream (Sparus aurata L.)
[J].
Estimation of genetic parameters for growth traits of Japanese flounder Paralichthys olivaceus using an animal mode
[J].
Maternal and genetic effects on individual variation during early development in Japanese flounder Paralichthys olivaceus
[J].

{kind=link}
{kind=link}