


中华锯齿米虾(Neocaridina denticulata sinensis)隶属节肢动物门(Arthropod)、甲壳纲(Crustacea)、十足目(Decapoda)、匙指虾科(Atyidae)、新米虾属(Neocaridina)[1],是亚洲的原生物种,广泛分布于河流、湖泊和水库中,因品系繁多、生存能力强、繁殖周期短、易于饲养等特性而深受水族爱好者的青睐。除具有观赏价值外,中华锯齿米虾还具有世代周期短、大型卵、成活率高等特点,是理想的实验和科研材料。野生中华锯齿米虾体色半透明、部分体色不透明、带有灰色树枝状条纹、少见红色和蓝色体色,而选育的中华锯齿米虾体色主要有红色、黄色、蓝色、橙色、透明色、黑色、琉璃色等色系。目前,有关中华锯齿米虾的研究[2⇓⇓⇓⇓⇓-8]主要集中在其生殖系统发育、性别分化、生理生化、毒理和胚胎发育及结构变化等方面,但关于其体色改良的研究则鲜见报道。
甲壳类动物虾青素代谢转化途径分为两大类:一类是将β-胡萝卜素转化为虾青素,如对虾类;另一类是直接利用中间产物叶黄素转化为虾青素,如龙虾和螃蟹[9-10]等。目前,已知类胡萝卜素超过600种,可分为分子中含氧原子的叶黄素类和不含氧原子、只含碳氢的胡萝卜素类两大类,其中类胡萝卜素的重要代表虾青素和叶黄素可作为色素添加剂而被广泛应用于动物饲料,以增加动物体色鲜艳度[11]。虾青素是甲壳类动物主要的呈色物质,但受提炼工艺不成熟且不易获取的影响,将其用于养殖活动会提高生产成本;而人工合成虾青素是通过化学方式合成的,产量高,成本低,但合成产物含有左旋(3S,3'S)、内消旋(3S,3'R)和右旋(3R,3'R)3种虾青素(同分异构体)[12]。螺旋藻的β-胡萝卜素含量为177.95 mg/100g,其被广泛应用于对虾饲料[13]。虽然大部分生物的体色主要由遗传物质控制,但是通过添加类胡萝卜素色素对体色产生的增艳效果可以提升其观赏和经济价值[14⇓⇓-17]。因此,本实验以红色、黄色和蓝色3个品系中华锯齿米虾为研究对象,根据薄层色谱分析结果挑选中华锯齿米虾的类胡萝卜素添加剂(螺旋藻粉、叶黄素、天然虾青素和人工合成虾青素),探究类胡萝卜素添加剂对其体色的影响,为提升工厂化养殖观赏虾类体色鲜艳度提供理论依据。
1 材料与方法
1.1 材料与仪器
实验用红色、黄色和蓝色3个品系中华锯齿米虾购于厦门本地的米虾养殖场,在实验室暂养14 d,暂养期间使用不含色素添加剂的饲料。
薄层色谱实验:β-胡萝卜素(标准品)和虾青素(标准品),上海甄准生物科技有限公司。P-20E型全自动薄层色谱点样仪,上海科哲生化科技有限公司;TK-20E全自动展开仪,上海科哲生化科技有限公司;Goodsee-10型薄层色谱成像仪、200 mm×200 mm薄层板,上海科哲生化科技有限公司。展层剂:石油醚:丙酮:正丙醇=90:10:0.45;0.1% BHT抗氧化剂:称取0.1 g BHT药品放入装有石油醚溶液的100 mL容量瓶中进行溶解,然后用石油醚定容,保存备用。
投喂实验:潘通色卡(PANTONE)为国际通用的标准色卡。天然虾青素和人工合成虾青素,长江水产研究所;螺旋藻粉,厦门嘉康饲料有限公司;叶黄素,广州立达尔生物科技股份有限公司;对虾配合饲料(不含色素饲料添加剂),厦门嘉康饲料有限公司。
1.2 薄层色谱分析
1.3 类胡萝卜素投喂实验
1)根据薄层色谱分析结果选择类胡萝卜素添加剂。
3)将中华锯齿米虾放入玻璃鱼缸(15 cm×20 cm×30 cm)饲养,分为2组排序编号,分别给予正常光照和黑暗环境,采用充气过滤循环系统。
4)实验开始前,在红色、黄色和蓝色3个品系中华锯齿米虾中,各随机抽取10只中华锯齿米虾进行体色比对。
5)实验设置:分别设置连续光照和黑暗条件。(1)光照组:光照强度为2 000 lx,红色、黄色和蓝色3个品系中华锯齿米虾分别设置4个实验组(分别添加天然虾青素、人工合成虾青素、叶黄素、螺旋藻粉)和1个对照组(不含饲料添加剂),每组设置3个平行,每个平行10只中华锯齿米虾。光照组共设置45个鱼缸。(2)黑暗组:红色、黄色和蓝色3个品系中华锯齿米虾分别设置4个实验组(分别添加天然虾青素、人工合成虾青素、叶黄素、螺旋藻粉)和1个对照组(不含饲料添加剂),每个组设置3个平行,每个平行10只中华锯齿米虾。黑暗组共设置45个鱼缸,鱼缸四周贴上不透光塑料板片,上方用遮阳网遮盖。
6)实验在室温25 ℃[水温(26±1)℃]的恒温条件下进行,pH 为7.0,氨氮为0 mg/L,亚硝酸盐为0 mg/L。周期为7 d,每天定时投喂1次,待中华锯齿米虾肠道饱满并离开摄食台后,开始吸污,并换水1/3。
7)观察指标:实验结束后,将各组中华锯齿米虾第二腹节的体色与潘通色卡进行比对,获得颜色型号,根据颜色型号找到相应的颜色组成,即透明度和子颜色,两者占比之和为100%。记录并分析色度值,并在解剖镜下分别观察光照和黑暗条件下对照组中华锯齿米虾体色的色素状态。
1.4 数据分析
为便于分析中华锯齿米虾体色的颜色型号,将Pro.Blue、ReF.Blue和Drak Blue的总和称为蓝色度值(Blue);Warm Red、Rub.Red、Red032和Bright Red的总和称为红色度值(Red);Yellow012和Yellow的总和称为黄色度值(Yellow);Orange021为橙色度值(Orange);其他颜色(Other)占比非常小,因而忽略不计。虾体色的颜色组成为:Trans.Wt+Blue+Red+Orange+Yellow+Other=100%,式中Trans.Wt为透明度值,该值越大,表示体色越浅。
实验结果采用SPSS 20.0进行单因素方差分析和多重比较。
2 结果与分析
2.1 薄层色谱分析中华锯齿米虾色素成分及饲料添加剂选择
薄层色谱分析结果显示,蓝色、红色、黄色3个品系中华锯齿米虾分别含有6个、5个、3个条带,并且都有相似的条带,说明3个品系中华锯齿米虾含有相同的色素成分。对比3个品系中华锯齿米虾与β-胡萝卜素、虾青素标准品的薄层色谱结果,发现3个品系中华锯齿米虾均具有2种标准品的条带,说明其均含有β-胡萝卜素和虾青素(图1)。因此选择含有β-胡萝卜素的螺旋藻和虾青素作为实验饲料色素添加剂。
图1
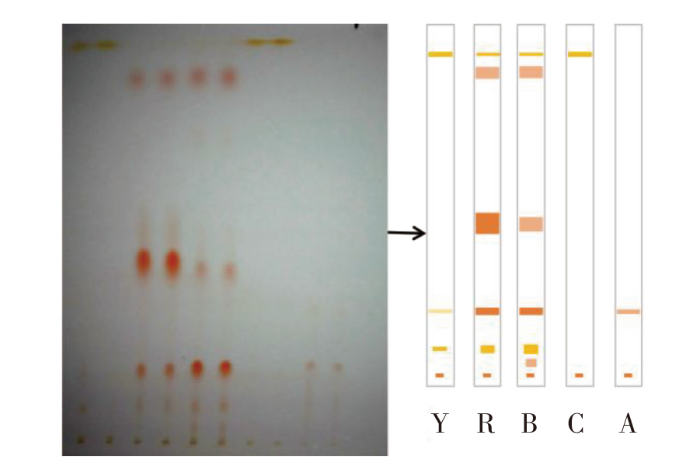
图1
中华锯齿米虾色素成分的薄层色谱结果
注:Y.黄色品系;R.红色品系;B.蓝色品系;C.β-胡萝卜素(标准品);A.虾青素(标准品)。自下而上第一个点为起始点。
Fig.1
TLC results of cytochrome component of N. denticulata sinensis
Notes:Y.The yellow strain;R.The red strain;B.The blue strain;C.β-carotene (standard);A.Astaxanthin(standard).The first point from bottom to top was the starting point.
2.2 连续光照条件下不同色素添加剂对中华锯齿米虾体色的影响
在连续光照条件下,3个品系中华锯齿米虾的存活率为100%,但3个品系的各实验组和对照组中华锯齿米虾的体色均发生了不同程度的变化。
在红色品系中:1)叶黄素组的中华锯齿米虾体色的透明度值(Trans.Wt)最低(13.19%),与实验前相比差异显著(P<0.05),与对照组和螺旋藻组相比无显著差异(P>0.05),与天然虾青素组和人工合成虾青素组相比差异显著(P<0.05)。2)螺旋藻组的中华锯齿米虾体色的红色值(Red)最大(54.87%),与实验前相比差异显著(P<0.05),与对照组、叶黄素组和天然虾青素组相比无显著差异(P>0.05),与人工合成虾青素组相比差异显著(P<0.05)。3)叶黄素组的中华锯齿米虾体色的黄色值(Yellow)最大(19.23%),与实验前、对照组、螺旋藻组和天然虾青素组相比无显著差异(P>0.05),与人工合成虾青素组相比差异显著(P<0.05)。4)各组中华锯齿米虾体色的橙色值(Orange)无显著差异(P>0.05)(图2)。
图2
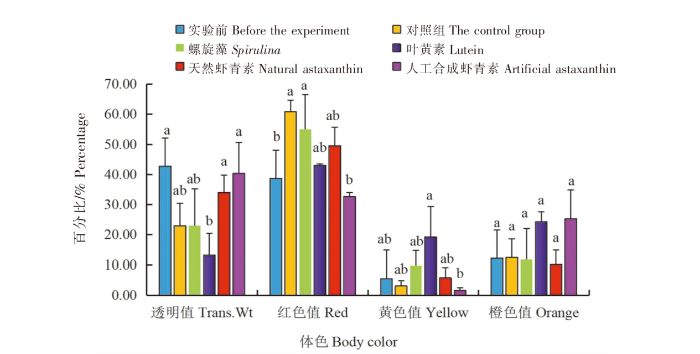
图2
光照条件下色素添加剂对红色品系中华锯齿米虾体色的影响
注:不同字母表示差异显著(P<0.05),相同字母无差异。以下同此。
Fig.2
Effect of pigment additives on the body color of the red strain of N. denticulata sinensis under light condition
Notes:Groups with different letters showed significant difference (P<0.05);groups with the same letter showed no difference.The same as below.
在黄色品系中:1)天然虾青素组中华锯齿米虾体色的透明度值(Trans.Wt)最低(31.17%),与实验前、对照组、螺旋藻组和人工合成虾青素组相比差异显著(P<0.05),与叶黄素组相比无显著差异(P>0.05)。2)各组中华锯齿米虾体色的红色值(Red)无显著差异(P>0.05)。3)天然虾青素组中华锯齿米虾体色的的黄色值(Yellow)最大(77.73 %),与实验前、对照组、螺旋藻组和人工合成虾青素组相比差异显著(P<0.05),与叶黄素组相比无显著差异(P>0.05)。4)各组中华锯齿米虾体色的橙色值(Orange)无显著差异(P>0.05)(图3)。
图3
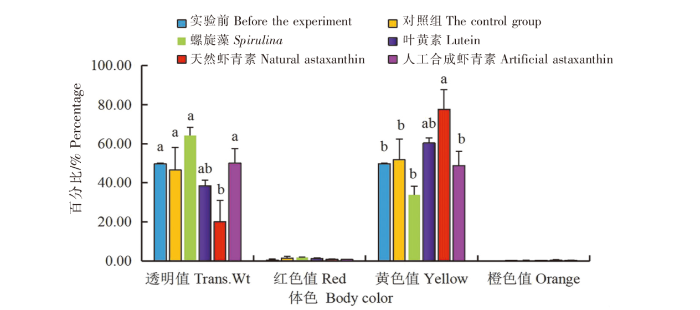
图3
光照条件下色素添加剂对黄色品系中华锯齿米虾体色的影响
Fig.3
Effect of pigment additives on the body color of the yellow strain of N. denticulata sinensis under light condition
在蓝色品系中:1)天然虾青素组中华锯齿米虾体色的透明度值(Trans.Wt)最低(53.02%),与实验前、对照组、螺旋藻组、叶黄素组和人工合成虾青素组相比差异显著(P<0.05)。2)各组中,中华锯齿米虾体色的红色值(Red)和黄色值(Yellow)均无显著差异(P>0.05)。3)天然虾青素组中华锯齿米虾体色的蓝色值(Blue)最大(77.73%),与实验前、对照组、螺旋藻组、叶黄素组和人工合成虾青素组相比差异显著(P<0.05)(图4)。
图4
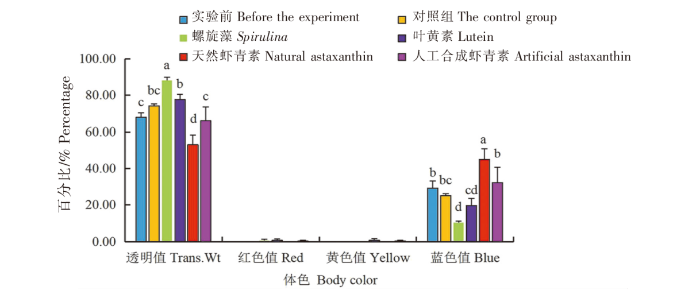
图4
光照条件下色素添加剂对蓝色品系中华锯齿米虾体色的影响
Fig.4
Effect of pigment additives on the body color of the blue strain of N. denticulata sinensis under light condition
以上结果显示,连续光照对红色品系中华锯齿米虾的红色素呈现出抑制效果。天然虾青素对黄色和蓝色2个品系中华锯齿米虾体色的促进效果最明显,与实验前相比分别提升29.56%和14.85%,与对照组相比分别提升26.31%和21.15%。
2.3 黑暗条件下不同色素添加剂对中华锯齿米虾体色的影响
在黑暗条件下,3个品系中华锯齿米虾的存活率为100%,其中部分实验组的体色发生不同程度的变化。
在红色品系中:1)各组中华锯齿米虾体色的透明度值(Trans.Wt)无显著差异(P>0.05)。2)人工合成虾青素组中华锯齿米虾体色的红色值(Red)最大(46.66%),与实验前相比无显著差异(P>0.05),与对照组、螺旋藻组相比差异显著(P<0.05),与叶黄素组和天然虾青素组相比无显著差异(P>0.05)。3)对照组中华锯齿米虾体色的黄色值(Yellow)最大(17.79%),与实验前和4个实验组相比差异显著(P<0.05),而实验前,4个实验组虾体之间的黄色值无显著差异(P>0.05)。4)实验前中华锯齿米虾体色的橙色值(Orange)最大(42.81%),螺旋藻组和叶黄素组的橙色值与对照组、天然虾青素组和人工合成虾青素组相比差异显著(P<0.05),天然虾青素组和人工合成虾青素组的橙色值与对照组之间无显著差异(P>0.05)(图5)。
图5
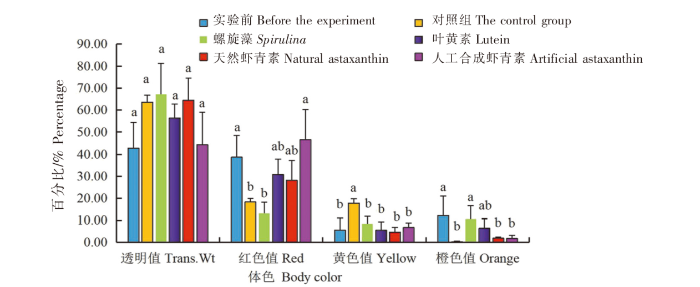
图5
黑暗条件下色素添加剂对红色品系中华锯齿米虾体色的影响
Fig.5
Effect of pigment additives on the body color of the red strain of N. denticulata sinensis under dark condition
在黄色品系中:1)实验前中华锯齿米虾体色的透明度值(Trans.Wt)最低(49.73%),与对照组和螺旋藻组相比差异显著(P<0.05),与叶黄素组、天然虾青素组和人工合成虾青素组相比无显著差异(P>0.05),对照组与4实验组之间无显著差异(P>0.05)。2)各组中华锯齿米虾体色的红色值(Red)无显著差异(P>0.05)。3)实验前中华锯齿米虾体色的黄色值(Yellow)最大(49.73%),与对照组和螺旋藻组相比差异显著(P<0.05),与叶黄素组、天然虾青素组和人工合成虾青素组相比无显著差异(P>0.05),对照组与4个实验组之间无显著差异(P>0.05)。4)各组中华锯齿米虾体色的橙色值(Orange)无显著差异(P>0.05)(图6)。
图6
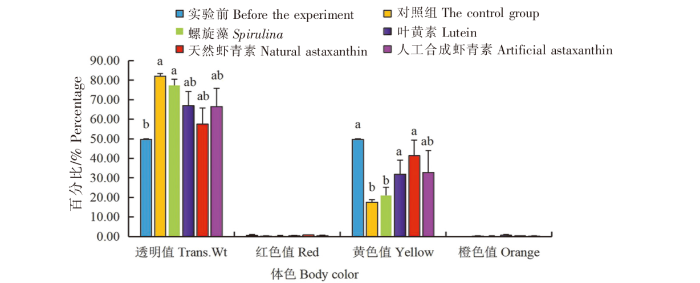
图6
黑暗条件下色素添加剂对黄色品系中华锯齿米虾体色的影响
Fig.6
Effect of pigment additives on the body color of the yellow strain of N. denticulata sinensis under dark condition
在蓝色品系中:1)实验前中华锯齿米虾体色的透明度值(Trans.Wt)最低(67.87%),与对照组、螺旋藻组、叶黄素组、天然虾青素组和人工合成虾青素组相比差异显著(P<0.05),对照组与4个实验组之间无显著差异(P>0.05)。2)各组中华锯齿米虾体色的红色值(Red)和黄色值(Yellow)无显著差异(P>0.05)。3)实验前中华锯齿米虾体色的蓝色值(Blue)最大(29.17%),与对照组、螺旋藻组、叶黄素组、天然虾青素组和人工合成虾青素组相比差异显著(P<0.05),对照组与4个实验组之间无显著差异(P>0.05)(图7)。
图7
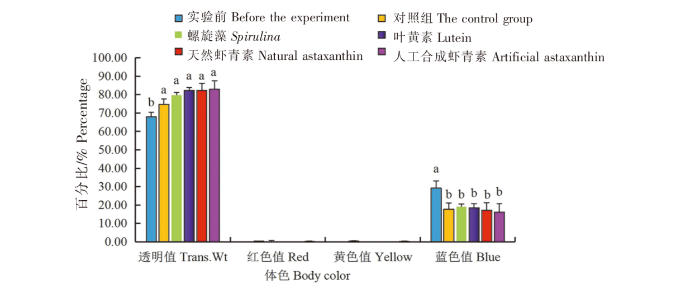
图7
黑暗条件下色素添加剂对蓝色品系中华锯齿米虾体色的影响
Fig.7
Effect of pigment additives on the body color of the blue strain of N. denticulata sinensis under dark condition
综上所述,在黑暗条件下,人工合成虾青素对红色品系中华锯齿米虾体色中的红色具有一定的维持效果;天然虾青素对黄色品系中华锯齿米虾体色的黄色具有一定的维持效果;4种色素添加剂对蓝色品系中华锯齿米虾的体色均不具有维持效果。
2.4 光照和黑暗条件下对照组中华锯齿米虾体色色素状态对比
图8
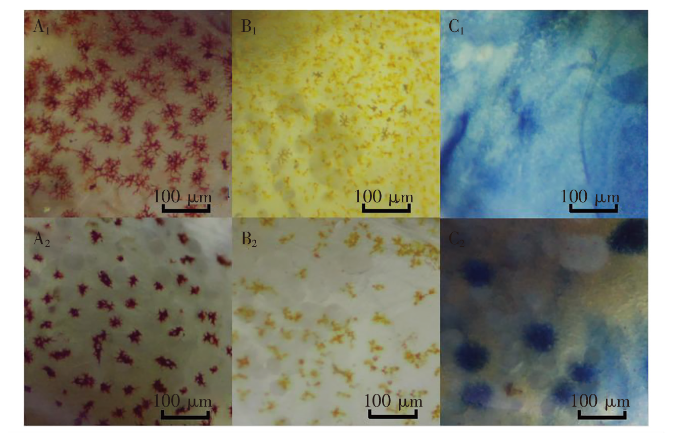
图8
实验结束后对照组中华锯齿米虾体色色素在不同条件下的状态
注:A.红色品系;B.黄色品系;C.蓝色品系;1.光照条件;2.黑暗条件。
Fig.8
After the experiment, the status of the pigment in the control group of N. denticulata sinensis under different conditions
Notes:A.The red strain;B.The yellow strain;C.The blue strain;1.Under light condition;2.Under dark condition.
3 讨论
3.1 中华锯齿米虾的色素成分差异
对红色、黄色和蓝色3个品系中华锯齿米虾的色素薄层色谱分析发现,3个品系都含有β-胡萝卜素和虾青素,但同时存在不同的条带,可能是类胡萝卜素的氧化代谢程度不同所致,说明不同品系对类胡萝卜素的氧化代谢程度有所差异[9]。
从薄层色谱分析结果可以推测,红色和黄色2个品系中华锯齿米虾体色的色素成分包含β-胡萝卜素和虾青素。而在对蓝色品系中华锯齿米虾进行色素提取时,发现加入石油醚后,其体色很快由蓝色变为红色。有研究指出,在蓝色龙虾中含有甲壳蓝蛋白,而甲壳蓝蛋白遇到石油醚会发生蛋白质变性而失活[26]。再结合薄层色谱分析结果,推测蓝色品系中华锯齿米虾体色的色素可能含有甲壳蓝蛋白与虾青素的结合物。因此,选择虾青素和富含β-胡萝卜素的螺旋藻作为饲料色素添加剂。而叶黄素作为β-胡萝卜素转化为虾青素的中间产物,其含量在生物体中相对较少,因而无法被薄层色谱分离并显示出来,但是作为类胡萝卜素的重要代表之一,其也能提升甲壳类动物的体色[9-10]。因此,本研究也同时将叶黄素作为饲料色素添加剂。
3.2 不同光照条件、不同类胡萝卜素对中华锯齿米虾体色的影响
本实验结果表明,在不同光照条件下,不同类胡萝卜素对不同品系的中华锯齿米虾体色的呈色效果表现出差异,其原因可能是不同品系的中华锯齿米虾对不同氧化级别的类胡萝卜素的响应有所不同,并且一定的光照能够促进黄色和蓝色品系中华锯齿米虾对类胡萝卜素的转化和利用。
在连续光照条件下,红色品系的中华锯齿米虾体色的透明度值(Trans.Wt)比实验前均有所降低,说明体色在一定程度有所增加;红色值(Red)结果显示,最大值为对照组,最小值为人工合成虾青素组,并且存在显著差异;部分实验组的黄色值(Yellow)和橙色值(Orange)与对照组和实验前相比都有一定程度的提升,但差异不显著。这可能有3种原因:1)从色素状态来看,长时间的光照会影响色素收缩,普通光连续照射可使红色素体发生色素聚集[27-28]。当色素体发生集聚时,红色素体外围会因红色素浓度降低而表现出橙色(图8:A1)。2)从色素合成上分析,在实验后期可能受到某些因素的抑制。类胡萝卜素的氧化途径位于线粒体内膜上,而在氧化过程中叶黄素形成的2,3-脱氢叶黄素和由玉米黄质形成的3,3’-二脱氢叶黄素是更有效的β-胡萝卜素双加氧酶2(BCDO2)氧化应激诱导剂,其会破坏线粒体内膜的氢离子(H+)平衡[29],因此为避免这两种物质造成的破坏,将其排除在线粒体外,或者进一步氧化,但该步骤需要损耗更多的氧。目前光动力效应致靶细胞线粒体损伤的机制研究表明,长期光照可使细胞内光敏剂血卟啉单甲醚(HMME)发生光漂白,导致HMME含量降低,线粒体在光敏化反应中产生较多的活性氧自由基,从而对其造成损伤[30]。因此,光照对红色品系的中华锯齿米虾的线粒体造成了一定的损伤,而为了满足机体的正常呼吸需求,线粒体将类胡萝卜素的大部分氧化途径关闭,造成了中间产物的颜色出现积累。3)在实验过程中,24 h连续光照使对照组的鱼缸出现藻类生长,作为底栖动物的中华锯齿米虾不停地抓食藻类而补充类胡萝卜素,导致对照组体色颜色加深。
在连续光照条件下,在黄色和蓝色2个品系中,天然虾青素组的中华锯齿米虾体色的透明度(Trans.Wt)均比实验前和对照组低,但其主要颜色黄色值(Yellow)和蓝色值(Blue)均比实验前和对照组高,说明天然虾青素对黄色和蓝色2个品系中华锯齿米虾体色具有显著的促进作用。研究表明,每克干鱼体质量含有的色素量与天然虾青素含量和投喂时间成正比[31-32],如以50 mg/kg虾青素饲料投喂缺乏虾青素的对虾4周后,其体色逐渐恢复至深青蓝色,并且还从虾壳中分离出类胡萝卜素[33];在不同的色素添加剂中表现出显著差异,说明黄色和蓝色2个品系中华锯齿米虾对不同色素添加剂的吸收利用率不同,如分别以虾青素和角黄素喂食虹鳟(Oncorhynchus mykiss),若要达到同样的着色效果,角黄素需72 mg/kg,而虾青素只需60 mg/kg,说明虾青素较角黄素的着色效率更高[34];投喂相同浓度的β-胡萝卜素、角黄素和虾青素,发现虾青素在组织中的积累量分别比角黄素和β-胡萝卜素高23%和43%[25]。在黄色和蓝色2个品系中华锯齿米虾中,人工合成虾青素组与天然虾青素组均表现出显著差异,这可能是由虾青素结构、功能和成分上的差异导致,虾青素具有左旋(3S,3'S)、内消旋(3S,3'R)和右旋(3R,3'R)3种同分异构体,而人工合成虾青素是以内消旋(3S,3'R)为主的3种结构的混合物,与养殖生物体内的虾青素以左旋(3S,3'S)为主截然不同[35];在生理功能方面,人工合成虾青素的稳定性和氧化活性均比天然虾青素差[36],并且在其合成过程中会产生其他化合物,从而降低其生物利用安全性[37]。
4 结论
1)3个品系的中华锯齿米虾含有的色素物质不同。红色、黄色和蓝色3个品系中华锯齿米虾的色素均含有β-类胡萝卜素和虾青素,蓝色品系的中华锯齿米虾色素可能含有甲壳蓝蛋白与虾青素的结合物。
2)在不同光照条件下,类胡萝卜素添加剂及对不同品系的中华锯齿米虾的体色影响存在差异。在光照条件下,红色品系的中华锯齿米虾体色的红色素受连续光照影响,导致色素出现聚集反应,使类胡萝卜素对其体色的促进效果不明显;在黄色和蓝色品系的中华锯齿米虾中,天然虾青素对其体色的促进效果最明显,与实验前相比分别提升29.56%和14.85%,与对照组相比分别提升26.31%和21.15%。在黑暗条件下,叶黄素、天然虾青素和人工合成虾青素对红色和黄色品系的中华锯齿米虾体色具有一定的促进、维持效果;4种类胡萝卜素添加剂对蓝色品系中华锯齿米虾体色均不具有维持效果。
参考文献
中华锯齿米虾卵母细胞卵黄发生的超微结构
[J].
利用透射电镜对中华锯齿米虾卵母细胞的卵黄发生过程进行超微结构的研究.结果表明:中华锯齿米虾卵黄发生过程是双源性的.在卵黄发生过程中,卵母细胞中的线粒体、内质网、溶酶体和核糖体等多种细胞器均参与了其内源性合成,其中线粒体最早参与形成卵黄颗粒.在卵黄发生的中后期,内质网成为内源性合成卵黄蛋白最主要的细胞器,并最终演化成颗粒较大且有膜包被的卵黄球;溶酶体也在此时期发挥其特有的吞噬功能,通过融合分解线粒体、内质网等胞质细胞器,最终形成一种十分致密的卵黄球.外源性卵黄则主要通过卵质膜形成的微绒毛和微吞饮小泡从卵周隙及滤泡细胞中摄取外源物质而形成.
中华锯齿米虾卵巢发育过程中卵黄蛋白积累的研究
[J].为了研究中华锯齿米虾(Neocaridina denticulata sinensis)卵巢发育过程中卵黄蛋白的积累情况,采用电泳洗脱法从其成熟卵巢中获得卵黄蛋白(卵黄磷蛋白,VN),制备多抗,进行双向免疫扩散和免疫印迹试验,结果显示,中华锯齿米虾的卵黄蛋白具有物种特异性和组织特异性,分别对孵化后12,32,43,55,70和82D的中华锯齿米虾进行解剖,免疫组织化学技术检测分析显示,仅82D的卵巢中出现免疫阳性反应,说明该阶段的卵巢中存在卵黄蛋白;用甲苯胺蓝染色分析同期结果显示与该结果一致本研究旨在了解中华锯齿米虾卵黄蛋白在卵巢发育过程中累积的时间节点,为中华锯齿米虾卵巢发育的研究提供基础资料。
A review of carotenoid utilisation and function in crustacean aquaculture
[J].
The biosynthesis of astaxanthin—ⅩⅣ.The conversion of labelled β-carotene-15,15'-3H2 into astaxanthin in the crab,Portunus trituberculatus
[J].
Pigmentation of prawn (Penaeus japonicus) with carotenoids
[J].
The molecular basis of the coloration mechanism in lobster shell:β-crustacyanin at 3.2 Åresolution
[J].\n The binding of the carotenoid astaxanthin (AXT) in the protein multimacromolecular complex crustacyanin (CR) is responsible for the blue coloration of lobster shell. The structural basis of the bathochromic shift mechanism has long been elusive. A change in color occurs from the orange red of the unbound dilute AXT (λ\n max\n 472 nm in hexane), the well-known color of cooked lobster, to slate blue in the protein-bound live lobster state (λ\n max\n 632 nm in CR). Intriguingly, extracted CR becomes red on dehydration and on rehydration goes back to blue. Recently, the innovative use of softer x-rays and xenon derivatization yielded the three-dimensional structure of the A\n 1\n apoprotein subunit of CR, confirming it as a member of the lipocalin superfamily. That work provided the molecular replacement search model for a crystal form of the β-CR holo complex, that is an A\n 1\n with A\n 3\n subunit assembly including two bound AXT molecules. We have thereby determined the structure of the A\n 3\n molecule\n de novo\n. Lobster has clearly evolved an intricate structural mechanism for the coloration of its shell using AXT and a bathochromic shift. Blue/purple AXT proteins are ubiquitous among invertebrate marine animals, particularly the Crustacea. The three-dimensional structure of β-CR has identified the protein contacts and structural alterations needed for the AXT color regulation mechanism.\n
Direct effects of visible and UVA light on pigment migration in erythrophores of nile tilapia
[J].Erythrophores derived from Nile tilapia (Oreochromis niloticus) are sensitive to visible light of defined wavelengths in primary culture in the same manner as erythrophores in the skin. Cultured erythrophores aggregate their pigment in response to light with peak wavelengths near 400 or 600 nm, while dispersion is caused by light near 500 nm. In this study, we report that ultraviolet A (UVA) with a peak wavelength near 365 nm also induces pigment aggregation in erythrophores in the skin and in primary culture. The responses of erythrophores in the skin or in culture depend on the light intensity, although the photo-sensitivity differs among individual cells. From the results, we conclude that the action of visible light and UVA light on tilapia erythrophores is direct, and that multiple types of visual pigments may coexist in individual erythrophores.
Is carotenoid ornamentation linked to the inner mitochondria membrane potential? A hypothesis for the maintenance of signal honesty
[J].Several mechanistic hypotheses have been proposed for how carotenoid pigmentation of integumentary structures can serve as an honest signal of individual quality. These hypotheses are founded on proposed links between carotenoids, immuno responsiveness, and oxidative stress, but an absence of biochemical information on the oxidative pathways of carotenoids has limited the sophistication of such hypotheses. Based on published evidence, we propose that the oxidation of carotenoids for the purpose of ornamentation in birds and reptiles is coupled to the inner mitochondria membrane. We predict that several carotenoid oxidation reactions yielding ornamental pigments occur on the inner mitochondrial membrane. Three of these reactions are proposed to occur within the ubiquinone biosynthesis cluster known as the Coq cluster consisting of approximately a dozen Coq members, tightly integrated and intimately associated with Complex I and III of the electron transport system. Ubiquinone and highly oxidized ornamental carotenoids share a stereochemically-conserved binding region suggesting that these two molecules may have shared similar pathways in the past. Carotenoids and ubiquinones may cooperate as redox participants in anti-radical reactions or independently in helping to maintain membrane or supra-complex stabilization during times of high-energy demand. Under this hypothesis, oxidation of carotenoids is coupled to the inner mitochondria membrane potential such that ornamental coloration reflects the efficiency of cellular respiration.Copyright © 2012 Elsevier Masson SAS. All rights reserved.
Correction of black tiger prawn (Penaeus monodon fabricius) coloration by astaxanthin
[J].
Digestibility of astaxanthin and canthaxanthin in rainbow trout as affected by dietary concentration,feeding rate and water salinity
[J].
Rapid liquid chromatographic method to distinguish wild salmon from aquacultured salmon fed synthetic astaxanthin
[J].Analytical methods are needed to determine the presence of color additives in fish. We report a liquid chromatographic (LC) method developed to identify the synthetic form of the color additive astaxanthin in salmon, based on differences in the relative ratios of the configurational isomers of astaxanthin. The distributions of configurational isomers of astaxanthin in the flesh of wild Atlantic and wild Pacific salmon are similar, but significantly different from that in aquacultured salmon. Astaxanthin is extracted from the flesh of salmon, passed through a silica gel Sep-Pak cartridge, and analyzed directly by LC on a Pirkle covalent L-leucine column. No derivatization of the astaxanthin is required-an important advantage of our approach, which is a modification of our previously described method. This method can be used to distinguish between aquacultured and wild salmon. The method has general applicability and can also be used to identify astaxanthins derived from other sources such as Phaffia yeast and Haematococcus pluvialis algae.
Separation of astaxanthin from red yeast Phaffia rhodozyma by supercritical carbon dioxide extraction
[J].
不同光源及光照时间对凡纳滨对虾(Litopenaeus vannamei)游离虾青素含量及生长的影响
[J].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}